Founder principle, Founder effect

創始者原理・創始者効果
Founder principle, Founder effect
解説:池田光穂
少数の個体(=創始者)たちが、もとの集団から分かれて新たな地域に進出したときに、その地域での創始者にな ることで、種分化の原動力になること。なぜ種分化の原動力になるのかというと、このような個体群(=集団)では、遺伝子浮動——ライト効果とも言い集団の 遺伝子頻度が世代間で偶発的に変動すること*——が起こりやすくなり、かつ新しい環境に適応した遺伝子給源(ジーン・プール)に淘汰がはたらく、つまり新 しい種の起源になりやすくなる。E.W.マイアーという進化学者が提唱した。
*任意に交配をおこなうN個体からなる有限集団では世代交代により配偶子が無作為に抽出されることにより遺伝子が変動し、その際の集団の平均ヘ テロ接合体の頻度は毎世代1/(2N)づつ減少する。つまりNが小さいと、その逆数に比例するヘテロ接合体の頻度の減少の割合は相対的に大きくなることが わかる。つまり創始者効果が現れやすくなるということだ。
| In population genetics, the founder effect
is the loss of genetic variation that occurs when a new population is
established by a very small number of individuals from a larger
population. It was first fully outlined by Ernst Mayr in 1942,[1] using
existing theoretical work by those such as Sewall Wright.[2] As a
result of the loss of genetic variation, the new population may be
distinctively different, both genotypically and phenotypically, from
the parent population from which it is derived. In extreme cases, the
founder effect is thought to lead to the speciation and subsequent
evolution of new species.[3] In the figure shown, the original population has nearly equal numbers of blue and red individuals. The three smaller founder populations show that one or the other color may predominate (founder effect), due to random sampling of the original population. A population bottleneck may also cause a founder effect, though it is not strictly a new population. The founder effect occurs when a small group of migrants—not genetically representative of the population from which they came—establish in a new area.[4][5] In addition to founder effects, the new population is often very small, so it shows increased sensitivity to genetic drift, an increase in inbreeding, and relatively low genetic variation. |
集団遺伝学において、創始者効果とは、大きな集団からごく少数の個体が
分かれて新たな集団が形成される際に起こる遺伝的多様性の損失を指す。1942年にエルンスト・マイヤーが初めて完全に概説し、[1]
セウォル・ライトなどの既存の理論的研究を引用した。[2]
遺伝的多様性の喪失の結果、新しい集団は、遺伝子型および表現型の両方において、派生元の親集団と著しく異なる場合がある。極端なケースでは、創始者効果
は種の分化と、それに続く新種の進化につながると考えられている。[3] 図に示されているように、元の集団では青と赤の個体がほぼ同数である。3つのより小さな創始者集団は、元の集団の無作為なサンプリングにより、いずれかの 色が優勢になる可能性があることを示している(創始者効果)。集団のボトルネックも厳密には新しい集団ではないが、創始者効果を引き起こす可能性がある。 創始者効果は、遺伝的に元の集団を代表していない小規模な移住集団が新しい地域に定住したときに起こる。[4][5] 創始者効果に加えて、新しい集団は非常に小規模であることが多いため、遺伝的浮動に対する感受性が高まり、近親交配が増加し、遺伝的多様性が比較的低いこ とが示される。 |
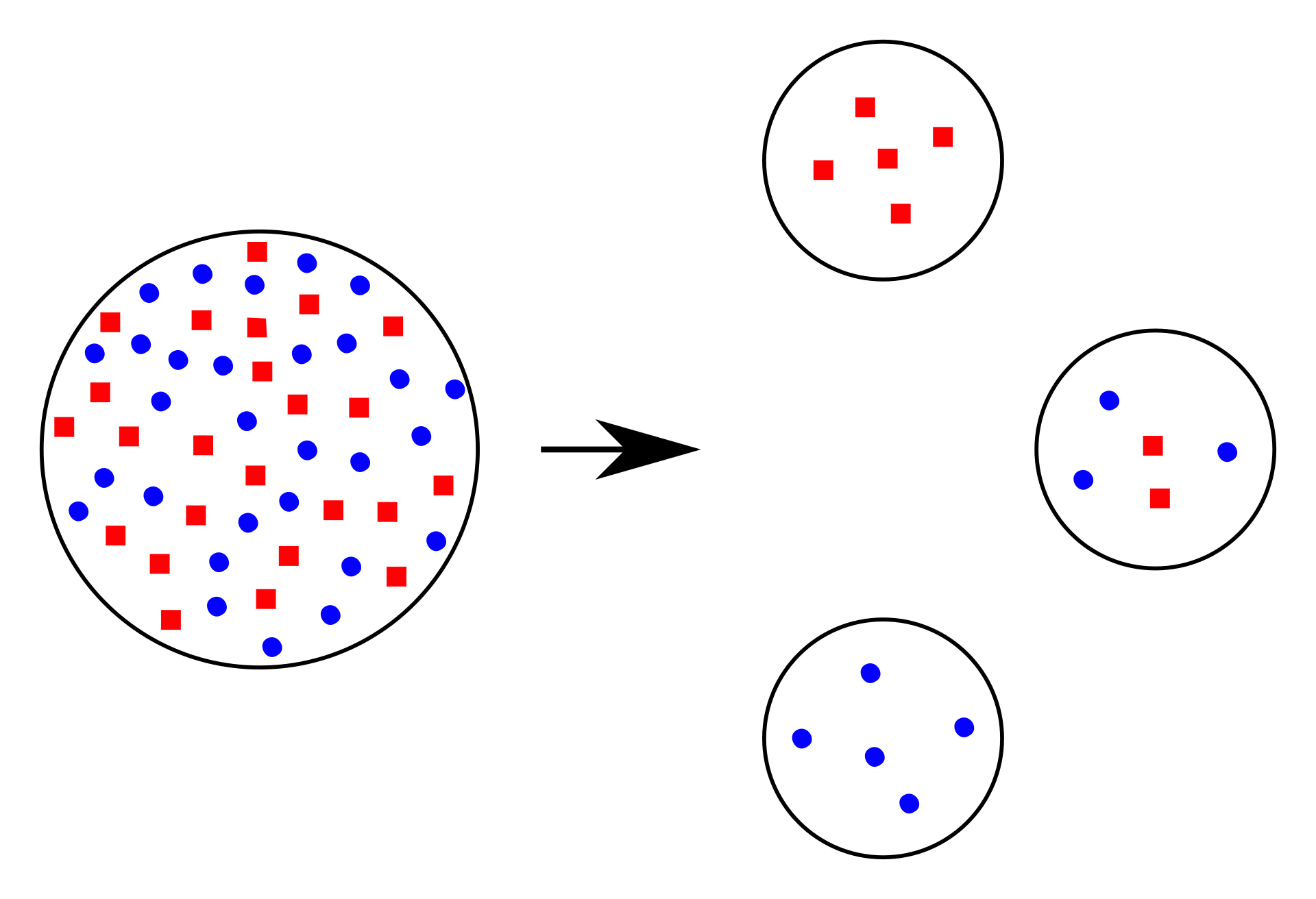 Founder effect: The original population (left) could give rise to different founder populations (right). |
創始者効果:元の集団(左)から、異なる創始者集団(右)が生まれる可能性がある。 |
| Founder mutation In genetics, a founder mutation is a mutation that appears in the DNA of one or more individuals which are founders of a distinct population. Founder mutations initiate with changes that occur in the DNA and can be passed down to other generations.[6][7] Any organism—from a simple virus to something complex like a mammal—whose progeny carry its mutation has the potential to express the founder effect,[8] for instance a goat[9][10] or a human.[11] Founder mutations originate in long stretches of DNA on a single chromosome; indeed, the original haplotype is the whole chromosome. As the generations progress, the proportion of the haplotype that is common to all carriers of the mutation is shortened (due to genetic recombination). This shortening allows scientists to roughly estimate the age of the mutation.[12] |
創始者変異 遺伝学において、創始者変異とは、特定の集団の創始者である1人または複数の個体のDNAに現れる変異である。創始者変異はDNAに生じる変化から始ま り、他の世代に受け継がれる可能性がある。[6][7] 単純なウイルスから哺乳類のような複雑な生物まで、その子孫が変異を継承するあらゆる生物は、創始者効果を発現する可能性がある。[8] 例えば、ヤギ[9][10]や人間[11]などである。 創始者変異は、単一の染色体上の長いDNA配列に由来する。実際、元のハプロタイプは染色体全体である。世代が進むにつれ、変異の保有者すべてに共通するハプロタイプの割合は短縮される(遺伝子組み換えによる)。この短縮により、科学者は変異の年代を概算することができる。[12] |
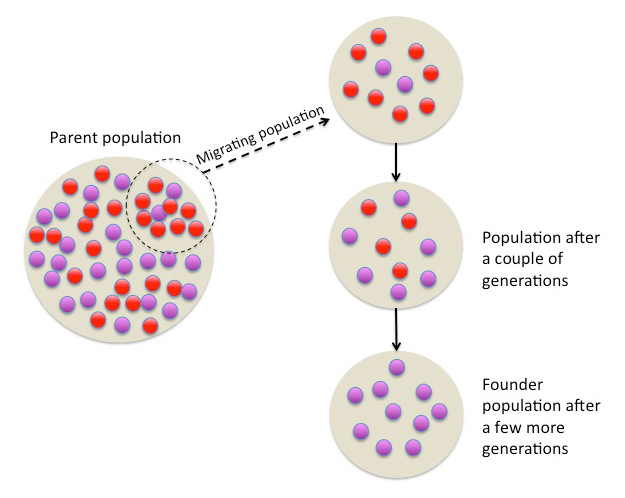 The small founding population experiences a loss of heterozygosity after multiple generations. (“Genetic Drift” by Boundless, 2015.[15]) |
小規模な創始者集団は、複数の世代を経てヘテロ接合性の喪失を経験する。(Boundless著『遺伝的浮動』(Genetic Drift)2015年。[15]) |
 The variation in gene frequency between the original population and colony may also trigger the two groups to diverge significantly over the course of many generations. As the variance, or genetic distance, increases, the two separated populations may become distinctively different, both genetically and phenotypically, although not only genetic drift, but also natural selection, gene flow and mutation all contribute to this divergence. This potential for relatively rapid changes in the colony's gene frequency led most scientists to consider the founder effect (and by extension, genetic drift) a significant driving force in the evolution of new species. Sewall Wright was the first to attach this significance to random drift and small, newly isolated populations with his shifting balance theory of speciation.[16] Following behind Wright, Ernst Mayr created many persuasive models to show that the decline in genetic variation and small population size accompanying the founder effect were critically important for new species to develop.[17] However, much less support for this view is shown today, since the hypothesis has been tested repeatedly through experimental research, and the results have been equivocal at best.[further explanation needed] Speciation by genetic drift is a specific case of peripatric speciation which in itself occurs in rare instances.[18] It takes place when a random change in genetic frequency of population favours the survival of a few organisms of the species with rare genes which cause reproductive mutation. These surviving organisms then breed among themselves over a long period of time to create a whole new species whose reproductive systems or behaviors are no longer compatible with the original population. [further explanation needed][19] |
元の集団とコロニーの遺伝子頻度の差異も、多くの世代を経るうちに2つの集団が著しく分岐するきっかけとなる可能性がある。分散、つまり遺伝的距離が大き くなると、2つの分離した集団は、遺伝的にも表現型的にも、はっきりと異なるものになる可能性がある。ただし、この分岐には、遺伝的浮動だけでなく、自然 淘汰、遺伝子流動、突然変異なども寄与する。集団の遺伝子頻度が比較的急速に変化する可能性があることから、ほとんどの科学者は、創始者効果(および、そ の延長線上にある遺伝的浮動)が、新種の進化における重要な推進力であると考えている。ソール・ライトは、種の分化に関するシフトバランス理論により、ラ ンダムなドリフトと新たに孤立した小規模な個体群にこのような重要性を初めて関連付けた。[16]ライトに続いて、エルンスト・マイヤーは、創始者効果に 伴う遺伝的多様性の減少と個体群の小規模化が、新種の進化にとって決定的に重要であることを示す説得力のあるモデルを数多く作成した。[17]しかし、今 日ではこの見解に対する支持は大幅に低下している。この仮説は実験的研究によって繰り返し検証されてきたが、結果はせいぜい曖昧なものでしかないためであ る。[さらなる説明が必要] 遺伝的浮動による種分化は、それ自体はまれなケースである周辺種分化の特殊なケースである。[18] それは、集団の遺伝頻度におけるランダムな変化が、生殖変異を引き起こすまれな遺伝子を持つ種の少数の生物の生存を助ける場合に起こる。こうして生き残っ た生物は、長い年月をかけて繁殖を繰り返し、生殖システムや行動様式がもはや元の個体群と適合しない全く新しい種を作り出す。[さらなる説明が必要] [19] |
| Serial founder effect Serial founder effects have occurred when populations migrate over long distances. Such long-distance migrations typically involve relatively rapid movements followed by periods of settlement. The populations in each migration carry only a subset of the genetic diversity carried from previous migrations. As a result, genetic differentiation tends to increase with geographic distance as described by the "isolation by distance" model.[20] The migration of humans out of Africa is characterized by serial founder effects.[21] Africa has the highest degree of human genetic diversity of any continent, which is consistent with an African origin of modern humans.[22] |
連続創始者効果 連続創始者効果は、個体群が長距離を移動する際に生じる。このような長距離移動は、通常、比較的速い移動に続いて定住期間がある。各移動における個体群 は、過去の移動から持ち込まれた遺伝的多様性の一部のみを運ぶ。その結果、地理的な距離が離れるにつれて遺伝的な分化が増加する傾向にある。これは「距離 による隔離」モデルによって説明されている。[20] 人類のアフリカ大陸外への移動は、連続した創始者効果によって特徴づけられる。[21] アフリカ大陸は、どの大陸よりも人類の遺伝的多様性が最も高い。これは現生人類がアフリカで誕生したという説と一致している。[22] |
| In island ecology Founder populations are essential to the study of island biogeography and island ecology. A natural "blank slate" is not easily found, but a classic series of studies on founder population effects was done following the catastrophic 1883 eruption of Krakatoa, which erased all life on the island.[23][24] Another continuing study has been following the biocolonization of Surtsey, Iceland, a new volcanic island that erupted offshore between 1963 and 1967. An earlier event, the Toba eruption in Sumatra about 73,000 years ago, covered some parts of India with 3–6 m (10–20 ft) of ash, and must have coated the Nicobar Islands and Andaman Islands, much nearer in the ash fallout cone, with life-smothering layers, forcing the restart of their biodiversity.[25] However, not all founder effect studies are initiated after a natural disaster; some scientists study the reinstatement of a species that became locally extinct or hadn't existed there before. A study has been in place since 1958 studying the wolf/moose interaction on Isle Royale in Lake Superior after those animals naturally migrated there, perhaps on winter ice. Hajji and others, and Hundertmark & Van Daele, studied the current population statuses of past founder effects in Corsican red deer and Alaskan elk, respectively. Corsican red deer are still listed as an endangered species, decades after a severe bottleneck. They inhabit the Tyrrhenian islands and surrounding mainlands currently, and before the bottleneck, but Hajji and others wanted to know how the deer originally got to the islands, and from what parent population or species they were derived. Through molecular analysis, they were able to determine a possible lineage, with red deer from the islands of Corsica and Sardinia being the most related to one another. These results are promising, as the island of Corsica was repopulated with red deer from the Sardinian island after the original Corsican red deer population became extinct, and the deer now inhabiting the island of Corsica are diverging from those inhabiting Sardinia.[26][27] Kolbe and others set up a pair of genetically sequenced and morphologically examined lizards on seven small islands to watch each new population's growth and adaptation to its new environment. Specifically, they were looking at the effects on limb length and perch width, both widely varying phenotypic ranges in the parent population. Unfortunately, immigration did occur, but the founder effect and adaptive differentiation, which could eventually lead to peripatric speciation, were statistically and biologically significant between the island populations after a few years. The authors also point out that although adaptive differentiation is significant, the differences between island populations best reflect the differences between founders and their genetic diversity that has been passed down through the generations.[28] Founder effects can affect complex traits, such as song diversity. In the Common Myna (Acridotheres tristis), the percentage of unique songs within a repertoire and within‐song complexity were significantly lower in birds from founder populations.[29] It was found by Tarr et al. (1998) that the loss of heterozygosity of the Laysan finch (Telespiza cantans) after founding events on small islands in the Pacific Ocean closely matched theoretical calculations upon examination of microsatellite loci.[30] |
島嶼生態学において、 創始者集団は島嶼生物地理学および島嶼生態学の研究に不可欠である。自然界で「白紙の状態」はなかなか見つからないが、1883年のクラカトアの壊滅的な 噴火により、島上の生物がすべて消滅したことを受けて、創始者集団の影響に関する一連の古典的研究が行われた。[23][24] また、1963年から1967年の間に沖合で噴火した新しい火山島であるアイスランドのサーツエイにおける生物の植民地化に関する研究も継続されている。 それより前の出来事としては、約7万3千年前にスマトラ島で起きたトバ火山の噴火では、インドの一部が3~6m(10~20フィート)の火山灰に覆われ、 さらに火山灰の降下円錐の中心に位置するニコバル諸島やアンダマン諸島は、生物を窒息させるほどの厚さの火山灰に覆われ、生物多様性の再出発を余儀なくさ れたに違いない。 しかし、創始者効果の研究がすべて自然災害後に開始されるわけではない。一部の科学者は、その地域で絶滅した種や、以前には存在していなかった種の復活に ついても研究している。1958年から、動物たちが自然に移動してきたと考えられる冬の氷上を移動して、スペリオル湖のロイヤル島でオオカミとヘラジカの 相互関係を研究している。ハジ氏と他の研究者たちは、コルシカのレッドディアにおける過去の孤立効果の現在の個体数状況を研究し、フンダーマーク氏とヴァ ン・ダール氏は、アラスカのヘラジカにおける孤立効果の現在の個体数状況を研究した。コルシカのレッドディアは、深刻な個体数減少の数十年後も、依然とし て絶滅危惧種としてリストアップされている。現在、彼らはティレニア海の島々および周辺の本土に生息しているが、ボトルネック以前にも生息していた。ハジ 氏らは、このシカがどのようにして島にたどり着いたのか、また、どの親個体群または種から派生したのかを知りたいと考えた。分子分析により、彼らは可能性 のある系統を特定することができた。コルシカ島とサルデーニャ島のシカが最も近縁である。コルシカ島では、元のコルシカ島の赤鹿の個体群が絶滅した後、サ ルデーニャ島の赤鹿が再導入されたが、現在コルシカ島に生息する鹿はサルデーニャ島に生息する鹿とは分化しつつある。この結果は有望である。[26] [27] コルベと他の研究者は、遺伝子配列と形態を調べた2匹のトカゲを7つの小島に放ち、それぞれの新しい個体群の成長と新しい環境への適応を観察した。具体的 には、親個体群において広く変異する表現型の範囲である肢の長さと止まり木の幅への影響を調べた。残念ながら、移住は起こったが、数年後には、最終的に周 辺種分化につながる可能性のある創始者効果と適応分化が、島々の個体群の間で統計的にも生物学上も有意な差となって現れた。著者らはまた、適応分化も重要 ではあるが、島々の個体群間の差異は、創始者間の差異と、世代を通じて受け継がれてきた遺伝的多様性の差異を最もよく反映していると指摘している。 創始者効果は、歌の多様性のような複雑な形質に影響を与える可能性がある。コモン・マイナ(Acridotheres tristis)では、レパートリー内の独特な歌の割合と、歌内の複雑性は、創始者集団の鳥では著しく低かった。 Tarr ら(1998年)は、太平洋の小島で起きた集団の孤立化の後にレイサンフィンチ(Telespiza cantans)でヘテロ接合性の喪失が起こったことを、マイクロサテライト遺伝子座の調査により理論計算とほぼ一致することを発見した。[30] |
| Among human populations Genetic studies of founder effect have concentrated on discovering ancestral and novel genetic diseases caused by founder effect and, to a lesser degree, on ancestry-related founder effects on populations, races, and ancient migrations, as well other aspects. The founder population could be the common ancestry of race or ethnicity or the forced localizations caused by artificial countries inside the larger group of ancestry, hence causing an original founder effect.[clarification needed] Race and specific founder effect mutation diseases are found in all races or ethnicities,[citation needed] and country-specific mutation diseases are caused by increasing homozygosity (the existence of same gene on both chromosomes pairs, hence a recessive disease may increase in just few generations). The genetic abnormality will increase incrementally with the decrease of number of isolated populations making tribe-specific diseases (such as Ashkenazis, Amish, and Bedouins) and novel genetic defects.[31] In recessive diseases, founder populations where underlying levels of genome-wide homozygosity are high due to shared common ancestry, but also for consanguineous populations that will have large genome-wide homozygous regions due to inbreeding. Having a catalog of disease-associated variation in these populations enables rapid, early, and accurate diagnoses that may improve patient outcomes due to informed clinical management and early interventions. Enclosed communities such as Amish communities, Ashkenazi communities, and relatively isolated islands allow scientists to better understand and further discover the mutated genes that cause these rare diseases and allow them to also discover protective genes as well.[32] Due to various migrations throughout human history, founder effects are somewhat common among humans in different times and places. The French Canadians of Quebec are a classical example of founder population. Over 150 years of French colonization, between 1608 and 1760, an estimated 8,500 pioneers married and left at least one descendant on the territory.[33] Following the takeover of the colony by the British crown in 1760, immigration from France effectively stopped, but descendants of French settlers continued to grow in number mainly due to their high fertility rate. Intermarriage occurred mostly with the deported Acadians and migrants coming from the British Isles. Since the 20th century, immigration in Quebec and mixing of French Canadians involve people from all over the world. While the French Canadians of Quebec today may be partly of other ancestries, the genetic contribution of the original French founders is predominant, explaining about 90% of regional gene pools, while Acadian (descended from other French settlers in eastern Canada) admixtures contributing 4% British and 2% Native American and other groups contributing less.[34] In humans, founder effects can arise from cultural isolation, and inevitably, endogamy. For example, the Amish populations in the United States exhibit founder effects because they have grown from a very few founders, have not recruited newcomers, and tend to marry within the community. Though still rare, phenomena such as polydactyly (extra fingers and toes, a symptom of a condition such as[35][36] Weyers acrodental dysostosis[35] or Ellis–Van Creveld syndrome[36]) are more common in Amish communities than in the American population at large.[37] Maple syrup urine disease affects about one out of 180,000 infants in the general population.[citation needed] Due in part to the founder effect,[38] however, the disease has a much higher prevalence in children of Amish, Mennonite, and Jewish descent.[39][40] Similarly, a high frequency of fumarase deficiency exists among the 10,000 members of the Fundamentalist Church of Jesus Christ of Latter Day Saints, a community which practices both endogamy and polygyny, where an estimated 75-80% of the community are blood relatives of just two men—founders John Y. Barlow and Joseph Smith Jessop.[41] In South Asia, castes like the Gujjars, the Baniyas and the Pattapu Kapu have estimated founder effects about 10 times as strong as those of Finns and Ashkenazi Jews.[42] In Africa, many members of the Vadoma tribe inherit ectrodactyly, giving them the nickname of the “two-toed tribe”. [43][failed verification] The island of Pingelap also suffered a population bottleneck in 1775 following a typhoon that had reduced the population to only 20 people. As a result, complete achromatopsia has a current rate of occurrence of roughly 10%, with an additional 30% being carriers of this recessive condition. Around 1814, a small group of British colonists founded a settlement on Tristan da Cunha, a group of small islands in the Atlantic Ocean, midway between Africa and South America. One of the early colonists apparently carried a rare, recessive allele for retinitis pigmentosa, a progressive form of blindness that afflicts homozygous individuals. As late as 1961, the majority of the genes in the gene pool on Tristan were still derived from 15 original ancestors; as a consequence of the inbreeding, of 232 people tested in 1961, four were suffering from retinitis pigmentosa. This represents a prevalence of 1 in 58, compared with a worldwide prevalence of around 1 in 4,000.[44] The abnormally high rate of twin births in Cândido Godói could be explained by the founder effect.[45] On 31 August 2023, researchers reported, based on genetic studies, that a human ancestor population bottleneck (from a possible 100,000 to 1000 individuals) occurred "around 930,000 and 813,000 years ago ... lasted for about 117,000 years and brought human ancestors close to extinction."[46][47] |
人類集団の間では、 創始者効果の遺伝学的研究は、創始者効果によって引き起こされる先祖代々受け継がれてきた遺伝病や新規の遺伝病の発見に集中しており、また、それほどでは ないが、集団、人種、古代の移住、その他の側面における先祖に関連する創始者効果にも焦点が当てられている。創始者集団は、人種や民族の共通の祖先である か、あるいは、より大きな祖先集団内の人工的な国によって強制的に局在化されたものであり、それゆえに、最初の創始者効果を引き起こした。人種や特定の創 始者効果変異疾患は、すべての人種や民族に見られる。国特有の変異疾患は、ホモ接合性の増加によって引き起こされる(同じ遺伝子が2組の染色体上に存在す ることであり、それゆえ劣性遺伝病はわずか数世代で増加する可能性がある)。隔離された集団の数が減少するにつれ、遺伝子異常は徐々に増加し、部族特有の 疾患(アシュケナージ、アーミッシュ、ベドウィンなど)や新たな遺伝的欠陥が生じる。劣性遺伝病では、共通の祖先を持つためゲノム全体のホモ接合性の基礎 レベルが高い創始者集団、および近親婚によりゲノム全体のホモ接合性の領域が大きい近親婚集団が存在する。これらの集団における疾患関連の変異のカタログ があれば、迅速かつ早期に正確な診断が可能となり、情報に基づいた臨床管理と早期介入により患者の予後を改善できる可能性がある。アーミッシュ社会、ア シュケナージ社会、比較的孤立した島嶼部などの閉鎖的な社会では、科学者たちがこれらの希少疾患の原因となる変異遺伝子をよりよく理解し、さらに発見する ことができ、また保護遺伝子も発見できる可能性がある。 人類の歴史におけるさまざまな移住により、創始者効果は、異なる時代や場所の人類の間でも、ある程度一般的である。ケベックのフランス系カナダ人は、創始 者集団の典型的な例である。1608年から1760年までの150年以上にわたるフランスによる植民地化の間に、推定8,500人の開拓者が結婚し、その 領土に少なくとも1人の子孫を残した。[33] 1760年にイギリス王冠が植民地を占領した後、フランスからの移民は事実上停止したが、フランス系入植者の子孫は、主に高い出生率により、その後も増加 し続けた。混血は主に追放されたアカディア人と、ブリテン諸島からの移民との間で行われた。20世紀以降、ケベックへの移民やフランス系カナダ人の混血 は、世界中の人々を巻き込むようになった。今日のケベックのフランス系カナダ人は、他の祖先を持つ者もいるが、元来のフランス人開拓者の遺伝的貢献が優勢 であり、この地域の遺伝子プールの約90%を占めている。一方、アカディア人(カナダ東部の他のフランス人入植者の子孫)の混血は、4%がイギリス人、 2%がネイティブアメリカンやその他のグループであり、それ以下である。 人間においては、創始者効果は文化的孤立や、必然的に同族婚から生じる可能性がある。例えば、米国のアーミッシュの人々は、創始者効果が認められる。なぜ なら、彼らはごく少数の創始者から成長し、新参者を勧誘せず、コミュニティ内で結婚する傾向があるからだ。まれではあるが、多指症(余分な指や足指、例え ば[35][36] ウェイアーズ先端骨異形成症[35] やエリス・ヴァン・クリーブランド症候群[36] などの症状)のような現象は、アメリカ社会全体よりもアーミッシュ社会においてより一般的である。[37] カエデシロップ尿症は、 。ただし、創始者効果の影響もあり、[38] アーミッシュ、メノナイト、ユダヤ人の子孫では、この病気の有病率がはるかに高い。[39][40] 同様に、末日聖徒イエス・キリスト教会原理主義派の1万人の信者には、高頻度でフマラーゼ欠損症が見られる。このコミュニティでは、 同教団では推定75~80%の信者が、創設者であるジョン・Y・バーロウとジョセフ・スミス・ジェサップの2人の男性の血縁者である。[41] 南アジアでは、グジャール族、バニヤ族、パッタプ・カプール族などのカーストにおいて、フィン人とアシュケナージ系ユダヤ人の10倍もの創始者効果がある と推定されている。[42] アフリカでは、ヴァドマ族の多くのメンバーが欠指症を遺伝しており、「二指族」というあだ名で呼ばれている。[43][検証失敗] ピンゲラップ島も1775年に台風により人口が20人にまで減少する人口の危機に直面した。その結果、完全色覚異常の発生率は現在およそ10%であり、さらに30%が劣性遺伝するこの状態の保因者である。 1814年頃、英国からの入植者たちの小集団が、アフリカと南アメリカの中間にある大西洋上の小さな島々、トリスタン・ダ・クーニャ諸島に定住した。初期 の入植者の一人が、網膜色素変性症のまれな劣性対立遺伝子を保有していたようだ。網膜色素変性症は、ホモ接合体の個体を冒す進行性の失明症である。 1961年になっても、トリスタン諸島の遺伝子プールの遺伝子の大半は、わずか15人の祖先から受け継がれたものであった。近親交配の結果、1961年に 検査を受けた232人のうち4人が網膜色素変性症を患っていた。これは、世界的な発生率が約4,000人に1人であるのに対し、58人に1人の割合であ る。 カンディド・ゴドイにおける双子の出生率が異常に高いのは、創始者効果によるものと考えられる。 2023年8月31日、研究者は遺伝学的研究に基づいて、人類の祖先が「約93万~81万3000年前に10万人から1000人へと人口が激減する集団の 危機に陥った」と報告した。この危機は「約11万7000年間続き、人類の祖先を絶滅の危機にまで追い込んだ」という。[46][47] |
| Cousin marriage Founder takes all Genetic bottleneck Genetic drift Inbreeding depression Mitochondrial Eve Neolithic founder crops Persister cells Popular sire effect Small population size |
近親婚 創始者一掃 遺伝的ボトルネック 遺伝的浮動 近交弱勢 ミトコンドリア・イブ 新石器時代の創始者作物 残留細胞 人気種牡馬効果 個体数の少なさ |
| https://en.wikipedia.org/wiki/Founder_effect |
|
| Genetic drift,
also known as random genetic drift, allelic drift or the Wright
effect,[1] is the change in the frequency of an existing gene variant
(allele) in a population due to random chance.[2] Genetic drift may cause gene variants to disappear completely and thereby reduce genetic variation.[3] It can also cause initially rare alleles to become much more frequent and even fixed. When few copies of an allele exist, the effect of genetic drift is more notable, and when many copies exist, the effect is less notable (due to the law of large numbers). In the middle of the 20th century, vigorous debates occurred over the relative importance of natural selection versus neutral processes, including genetic drift. Ronald Fisher, who explained natural selection using Mendelian genetics,[4] held the view that genetic drift plays at most a minor role in evolution, and this remained the dominant view for several decades. In 1968, population geneticist Motoo Kimura rekindled the debate with his neutral theory of molecular evolution, which claims that most instances where a genetic change spreads across a population (although not necessarily changes in phenotypes) are caused by genetic drift acting on neutral mutations.[5][6] In the 1990s, constructive neutral evolution was proposed which seeks to explain how complex systems emerge through neutral transitions.[7][8] Analogy with marbles in a jar The process of genetic drift can be illustrated using 20 marbles in a jar to represent 20 organisms in a population.[9] Consider this jar of marbles as the starting population. Half of the marbles in the jar are red and half are blue, with each colour corresponding to a different allele of one gene in the population. In each new generation, the organisms reproduce at random. To represent this reproduction, randomly select a marble from the original jar and deposit a new marble with the same colour into a new jar. This is the "offspring" of the original marble, meaning that the original marble remains in its jar. Repeat this process until 20 new marbles are in the second jar. The second jar will now contain 20 "offspring", or marbles of various colours. Unless the second jar contains exactly 10 red marbles and 10 blue marbles, a random shift has occurred in the allele frequencies. If this process is repeated a number of times, the numbers of red and blue marbles picked each generation fluctuates. Sometimes, a jar has more red marbles than its "parent" jar and sometimes more blue. This fluctuation is analogous to genetic drift – a change in the population's allele frequency resulting from a random variation in the distribution of alleles from one generation to the next. In any one generation, no marbles of a particular colour could be chosen, meaning they have no offspring. In this example, if no red marbles are selected, the jar representing the new generation contains only blue offspring. If this happens, the red allele has been lost permanently in the population, while the remaining blue allele has become fixed: all future generations are entirely blue. In small populations, fixation can occur in just a few generations. |
遺伝的浮動(ランダム遺伝的浮動、対立遺伝子浮動、ライト効果とも呼ばれる)とは、[1] 集団内の既存の遺伝子変異体(対立遺伝子)の頻度が、偶然によって変化することである。[2] 遺伝的浮動は、遺伝子変異体が完全に消失し、遺伝的多様性が減少する原因となる可能性がある。[3] また、当初はまれな対立遺伝子が、はるかに頻繁に現れるようになり、固定される原因となる可能性もある。 対立遺伝子のコピー数が少ない場合、遺伝的浮動の影響はより顕著になり、コピー数が多い場合、その影響はより顕著にはならない(大数の法則による)。20 世紀中頃には、自然淘汰とニュートン過程(遺伝的浮動を含む)の相対的重要性について活発な議論が交わされた。メンデル遺伝学を用いて自然淘汰を説明した ロナルド・フィッシャーは、遺伝的浮動は進化においてせいぜい補助的な役割しか果たさないという見解を示し、この見解は数十年にわたって支配的であった。 1968年、集団遺伝学者の木村資生は、分子進化の中立理論を提唱し、この議論を再燃させた。この理論では、集団内で遺伝的変化が拡散する(必ずしも表現 型の変化とは限らない)ほとんどの事例は、中立的な突然変異に作用する遺伝的浮動によって引き起こされると主張している。[5][6] 1990年代には、中立進化説を発展させた「構成的中立進化」が提唱され、中立的な変遷を通じて複雑なシステムがどのようにして出現するのかを説明しよう としている。[7][8] 瓶の中のビー玉に例える 遺伝的浮動のプロセスは、瓶の中のビー玉20個で集団内の20個の生物を表すことで説明できる。[9] このビー玉の瓶を初期の集団と考える。瓶の中のビー玉は半分が赤、半分が青で、それぞれの色が集団内の異なる遺伝子の対立遺伝子に対応している。各世代で は、生物はランダムに繁殖する。この繁殖を表現するために、元の瓶からランダムにビー玉を1つ選び、同じ色の新しいビー玉を新しい瓶に入れる。これは元の ビー玉の「子孫」であり、元のビー玉は元の瓶に残っている。このプロセスを繰り返し、20個の新しいビー玉が2番目の瓶に入るまで続ける。2番目の瓶に は、20個の「子孫」、つまりさまざまな色のビー玉が入っている。2番目の瓶に赤いビー玉がちょうど10個と青いビー玉がちょうど10個入っていない限 り、対立遺伝子の頻度にランダムな変化が生じている。 このプロセスを何度も繰り返すと、各世代で選ばれる赤と青のビー玉の数が変動する。ある瓶には「親」の瓶よりも赤いビー玉が多く入っていることもあれば、 青いビー玉が多いこともある。この変動は遺伝的浮動に類似している。すなわち、世代から世代へと、対立遺伝子の分布がランダムに変化することで集団の対立 遺伝子頻度が変化することである。 ある世代では、特定の色のビー玉が選ばれることはなく、つまり、その色のビー玉には子孫がいないことになる。この例で、赤いビー玉が選ばれなかった場合、 新世代を表す瓶には青いビー玉の子孫しか入っていない。このようなことが起こると、赤い対立遺伝子は集団から永久に失われ、残った青い対立遺伝子は固定さ れる。つまり、将来の世代はすべて青一色になる。集団が小さい場合、固定はわずか数世代で起こりうる。 |
In this simulation, each black
dot on a marble signifies that it has been chosen for copying
(reproduction) one time. Fixation in the blue "allele" occurs within
five generations.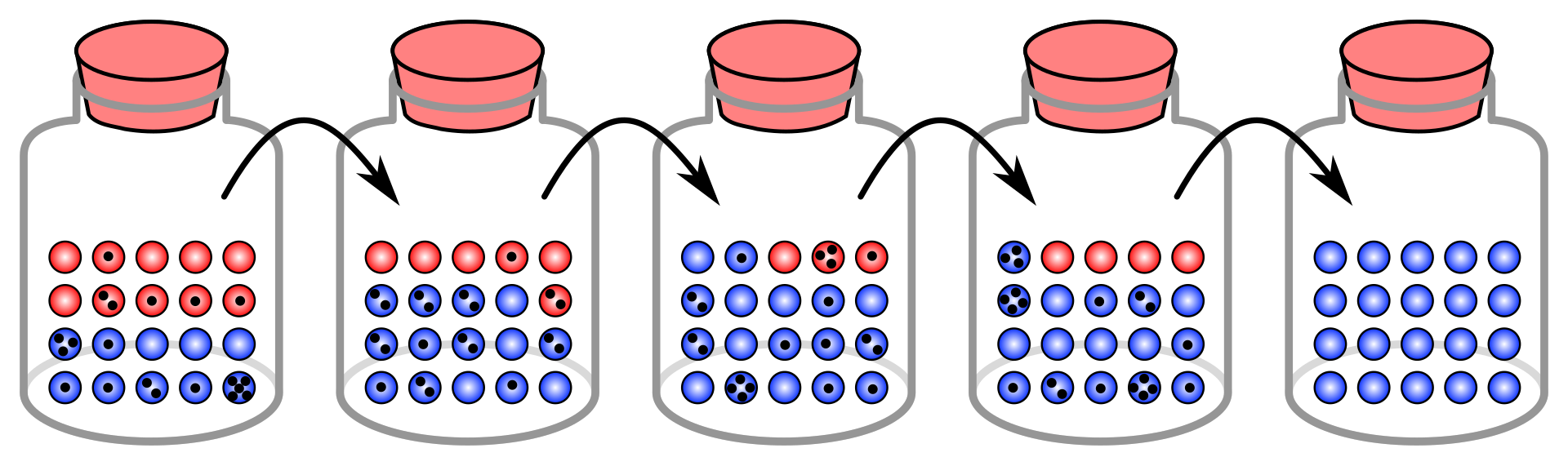 |
このシミュレーションでは、ビー玉上の黒い点は、1回コピー(再生)されたことを意味する。青い「対立遺伝子」の固定は5世代以内で起こる。 |
| https://en.wikipedia.org/wiki/Genetic_drift |
|
| Versus natural selection In natural populations, genetic drift and natural selection do not act in isolation; both phenomena are always at play, together with mutation and migration. Neutral evolution is the product of both mutation and drift, not of drift alone. Similarly, even when selection overwhelms genetic drift, it can only act on variation that mutation provides. While natural selection has a direction, guiding evolution towards heritable adaptations to the current environment, genetic drift has no direction and is guided only by the mathematics of chance.[29] As a result, drift acts upon the genotypic frequencies within a population without regard to their phenotypic effects. In contrast, selection favors the spread of alleles whose phenotypic effects increase survival and/or reproduction of their carriers, lowers the frequencies of alleles that cause unfavorable traits, and ignores those that are neutral.[30] The law of large numbers predicts that when the absolute number of copies of the allele is small (e.g., in small populations), the magnitude of drift on allele frequencies per generation is larger. The magnitude of drift is large enough to overwhelm selection at any allele frequency when the selection coefficient is less than 1 divided by the effective population size. Non-adaptive evolution resulting from the product of mutation and genetic drift is therefore considered to be a consequential mechanism of evolutionary change primarily within small, isolated populations.[31] The mathematics of genetic drift depend on the effective population size, but it is not clear how this is related to the actual number of individuals in a population.[17] Genetic linkage to other genes that are under selection can reduce the effective population size experienced by a neutral allele. With a higher recombination rate, linkage decreases and with it this local effect on effective population size.[32][33] This effect is visible in molecular data as a correlation between local recombination rate and genetic diversity,[34] and negative correlation between gene density and diversity at noncoding DNA regions.[35] Stochasticity associated with linkage to other genes that are under selection is not the same as sampling error, and is sometimes known as genetic draft in order to distinguish it from genetic drift.[17] Low allele frequency makes alleles more vulnerable to being eliminated by random chance, even overriding the influence of natural selection. For example, while disadvantageous mutations are usually eliminated quickly within the population, new advantageous mutations are almost as vulnerable to loss through genetic drift as are neutral mutations. Not until the allele frequency for the advantageous mutation reaches a certain threshold will genetic drift have no effect.[30] |
自然淘汰に対する 自然集団において、遺伝的浮動と自然淘汰は単独で作用するのではなく、両方の現象は常に突然変異や移動とともに行われている。中立進化は突然変異と浮動の 両方の産物であり、浮動のみの産物ではない。同様に、選択が遺伝的浮動を圧倒する場合でも、突然変異がもたらす変化に対してのみ作用する。 自然淘汰には方向性があり、進化を現在の環境への遺伝的適応へと導くが、遺伝的浮動には方向性がなく、偶然の数学のみによって導かれる。[29] その結果、浮動は表現型の影響を考慮することなく、集団内の遺伝子型の頻度に作用する。これに対し、選択は表現型の影響がその保有者の生存率や繁殖率を高 める対立遺伝子の拡散を促し、好ましくない形質を引き起こす対立遺伝子の頻度を低下させ、中立的なものは無視する。 大数の法則によれば、対立遺伝子のコピーの絶対数が少ない場合(例えば、小規模な集団の場合)、世代ごとの対立遺伝子頻度のドリフトの大きさは大きくな る。ドリフトの大きさは、選択係数が1を実効集団サイズで割った値よりも小さい場合、どの対立遺伝子頻度においても選択を圧倒するほど大きい。突然変異と 遺伝的浮動の産物である非適応進化は、主に小規模で孤立した集団における進化の変化の必然的なメカニズムであると考えられている。[31] 遺伝的浮動の数学は実効集団サイズに依存するが、これが集団内の実際の個体数とどのように関連しているかは明らかではない。[17] 選択を受けている他の遺伝子との遺伝的連鎖は、中立対立遺伝子が経験する実効集団サイズを減少させる可能性がある。組み換え率が高くなると連鎖は弱まり、 それに伴って有効集団サイズに対する局所的な影響も弱まる。[32][33] この影響は、局所的な組み換え率と遺伝的多様性との相関関係として分子データに現れるほか、 非コードDNA領域における遺伝子密度と多様性との間には負の相関がある。[35] 選択圧を受けている他の遺伝子との連鎖に関連する確率性はサンプリングエラーとは異なり、遺伝的浮動との区別のために遺伝的草案と呼ばれることもある。 [17] 対立遺伝子の頻度が低いと、自然淘汰の影響を覆すことさえあり、ランダムな偶然によって排除されやすくなる。例えば、不利な突然変異は通常、集団内で速や かに排除されるが、新しい有利な突然変異は、中立突然変異と同様に、遺伝的浮動によって失われる可能性がほぼ同じである。有利な突然変異の対立遺伝子頻度 が一定の閾値に達するまでは、遺伝的浮動の影響はない。[30] |
| Population bottleneck Main article: Population bottleneck 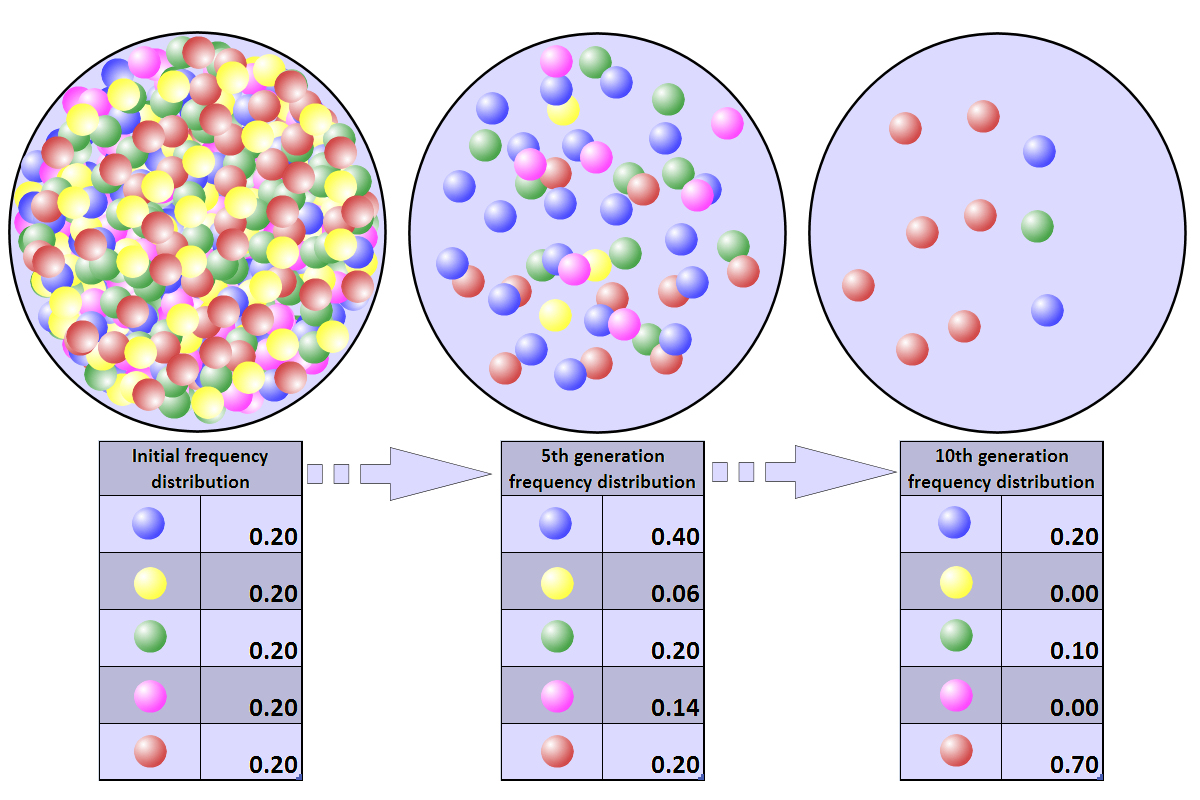 Changes in a population's allele frequency following a population bottleneck: the rapid and radical decline in population size has reduced the population's genetic variation. A population bottleneck is when a population contracts to a significantly smaller size over a short period of time due to some random environmental event. In a true population bottleneck, the odds for survival of any member of the population are purely random, and are not improved by any particular inherent genetic advantage. The bottleneck can result in radical changes in allele frequencies, completely independent of selection.[36] The impact of a population bottleneck can be sustained, even when the bottleneck is caused by a one-time event such as a natural catastrophe. An interesting example of a bottleneck causing unusual genetic distribution is the relatively high proportion of individuals with total rod cell color blindness (achromatopsia) on Pingelap atoll in Micronesia.[37] After a bottleneck, inbreeding increases. This increases the damage done by recessive deleterious mutations, in a process known as inbreeding depression. The worst of these mutations are selected against, leading to the loss of other alleles that are genetically linked to them, in a process of background selection.[2] For recessive harmful mutations, this selection can be enhanced as a consequence of the bottleneck, due to genetic purging. This leads to a further loss of genetic diversity. In addition, a sustained reduction in population size increases the likelihood of further allele fluctuations from drift in generations to come. A population's genetic variation can be greatly reduced by a bottleneck, and even beneficial adaptations may be permanently eliminated.[38] The loss of variation leaves the surviving population vulnerable to any new selection pressures such as disease, climatic change or shift in the available food source, because adapting in response to environmental changes requires sufficient genetic variation in the population for natural selection to take place.[39][40] There have been many known cases of population bottleneck in the recent past. Prior to the arrival of Europeans, North American prairies were habitat for millions of greater prairie chickens. In Illinois alone, their numbers plummeted from about 100 million birds in 1900 to about 50 birds in the 1990s. The declines in population resulted from hunting and habitat destruction, but a consequence has been a loss of most of the species' genetic diversity. DNA analysis comparing birds from the mid century to birds in the 1990s documents a steep decline in the genetic variation in just the latter few decades. Currently the greater prairie chicken is experiencing low reproductive success.[41] However, the genetic loss caused by bottleneck and genetic drift can increase fitness, as in Ehrlichia.[42] Over-hunting also caused a severe population bottleneck in the northern elephant seal in the 19th century. Their resulting decline in genetic variation can be deduced by comparing it to that of the southern elephant seal, which were not so aggressively hunted.[43] Founder effect Main article: Founder effect 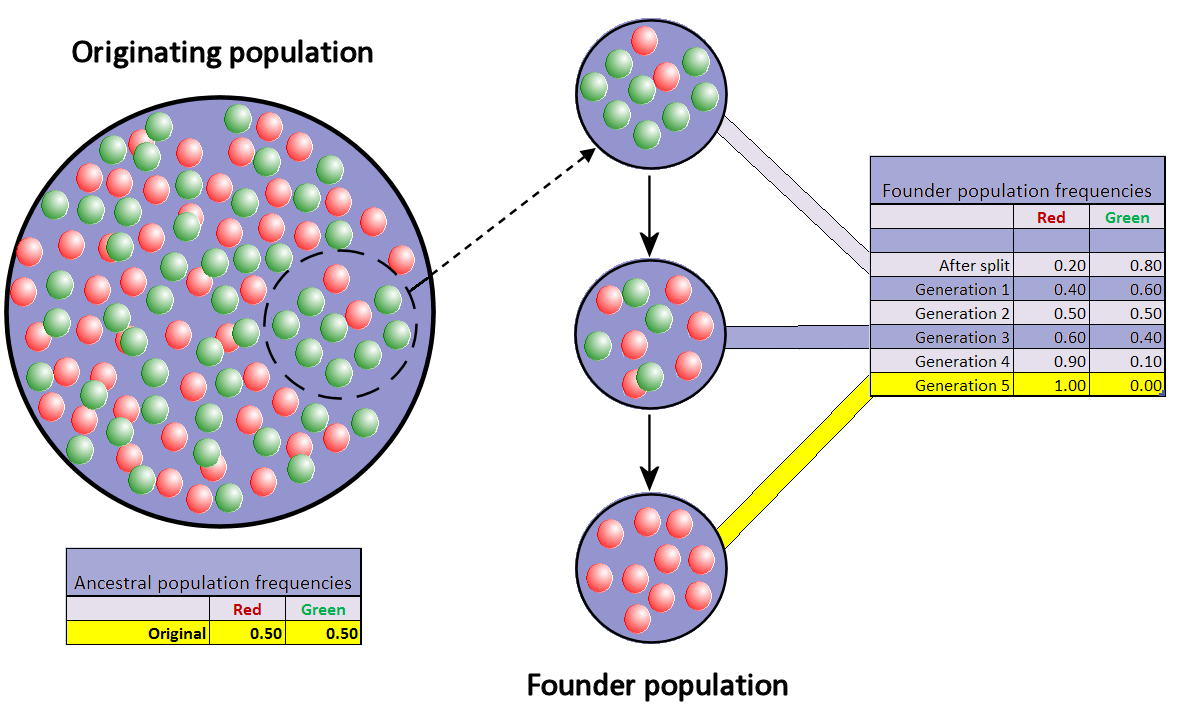 When very few members of a population migrate to form a separate new population, the founder effect occurs. For a period after the foundation, the small population experiences intensive drift. In the figure this results in fixation of the red allele. The founder effect is a special case of a population bottleneck, occurring when a small group in a population splinters off from the original population and forms a new one. The random sample of alleles in the just formed new colony is expected to grossly misrepresent the original population in at least some respects.[44] It is even possible that the number of alleles for some genes in the original population is larger than the number of gene copies in the founders, making complete representation impossible. When a newly formed colony is small, its founders can strongly affect the population's genetic make-up far into the future. A well-documented example is found in the Amish migration to Pennsylvania in 1744. Two members of the new colony shared the recessive allele for Ellis–Van Creveld syndrome. Members of the colony and their descendants tend to be religious isolates and remain relatively insular. As a result of many generations of inbreeding, Ellis–Van Creveld syndrome is now much more prevalent among the Amish than in the general population.[30][45] The difference in gene frequencies between the original population and colony may also trigger the two groups to diverge significantly over the course of many generations. As the difference, or genetic distance, increases, the two separated populations may become distinct, both genetically and phenetically, although not only genetic drift but also natural selection, gene flow, and mutation contribute to this divergence. This potential for relatively rapid changes in the colony's gene frequency led most scientists to consider the founder effect (and by extension, genetic drift) a significant driving force in the evolution of new species. Sewall Wright was the first to attach this significance to random drift and small, newly isolated populations with his shifting balance theory of speciation.[46] Following after Wright, Ernst Mayr created many persuasive models to show that the decline in genetic variation and small population size following the founder effect were critically important for new species to develop.[47] However, there is much less support for this view today since the hypothesis has been tested repeatedly through experimental research and the results have been equivocal at best.[48] |
人口のボトルネック 詳細は「人口のボトルネック」を参照 人口のボトルネックに続く集団の対立遺伝子頻度の変化:個体数の急速かつ急激な減少により、集団の遺伝的多様性が減少した。 個体群の急激な縮小とは、何らかの突発的な環境要因により、個体群が短期間に著しく小さなサイズに縮小することである。真の個体群の急激な縮小では、個体 群のどの個体が生き残るかという確率は完全にランダムであり、特定の遺伝的優位性によって改善されることはない。急激な縮小は、選択とは全く無関係に、対 立遺伝子頻度の急激な変化をもたらす可能性がある。 集団のボトルネックの影響は、それが自然災害のような一度限りの出来事によって引き起こされた場合でも持続する可能性がある。 ボトルネックが特異な遺伝子分布を引き起こす興味深い例としては、ミクロネシアのピンゲラップ環礁における杆体細胞完全色盲(無色覚)の個体の割合が比較 的高いことが挙げられる。[37] ボトルネックの後には近親交配が増加する。これにより劣性有害突然変異による被害が増加し、同系交配弱勢と呼ばれる過程が起こる。これらの突然変異のうち 最悪のものは淘汰され、背景選択の過程で遺伝的に関連する他の対立遺伝子を失うことになる。劣性有害突然変異の場合、この淘汰は遺伝的浄化の結果として、 ボトルネックの結果として強化される可能性がある。これにより、遺伝的多様性がさらに失われることになる。さらに、個体数の持続的な減少は、次世代以降の 世代における対立遺伝子の変動の可能性を高める。 個体群の遺伝的多様性は、ボトルネックによって大幅に減少する可能性があり、有益な適応さえも恒久的に排除される可能性がある。[38] 変動の喪失は、病気、気候変動、利用可能な食料源の変化など、新たな選択圧に対して生存する個体群を脆弱にする。なぜなら、環境の変化に対応する適応に は、自然淘汰が起こるために個体群に十分な遺伝的多様性が必要だからである。[39][40] 過去にも、個体群のボトルネック現象が数多く知られている。ヨーロッパ人が到着する以前、北米の草原には何百万羽もの草原雷鳥が生息していた。イリノイ州 だけでも、1900年には約1億羽いた個体数が、1990年代には約50羽にまで激減した。個体数の減少は狩猟と生息地の破壊によるものだが、その結果、 種の遺伝的多様性のほとんどが失われた。1950年代の鳥と1990年代の鳥を比較したDNA分析では、わずか後者の数十年間で遺伝的多様性が急激に減少 したことが記録されている。現在、オオライチョウは繁殖成功率が低い状態にある。[41] しかし、ボトルネックや遺伝的浮動によって引き起こされた遺伝子の損失は、エリキア属のように適応度を高める可能性がある。[42] 19世紀には、過剰な狩猟により、ミナミゾウアザラシに深刻な個体数のボトルネックが生じた。その結果、遺伝的多様性が低下したことは、それほど狩猟の対象とならなかったミナミゾウアザラシと比較することで推測できる。[43] 創始者効果 詳細は「創始者効果」を参照 個体群の非常に少数の個体が移住して別の個体群を形成する場合、創始者効果が起こる。創始後の一定期間、その小規模な個体群は集中的なドリフトを経験する。図では、これが赤の対立遺伝子の固定につながる。 創始者効果は、個体群の小集団が元の個体群から分裂して新たな個体群を形成する際に起こる、個体群のボトルネックの特殊なケースである。形成されたばかり の新しい集団における対立遺伝子のランダムサンプルは、少なくともいくつかの点において、元の集団を大幅に誤って表現することが予想される。[44] 元の集団におけるいくつかの遺伝子に対する対立遺伝子の数が、創始者集団における遺伝子コピーの数よりも多い場合もあり、完全な表現が不可能になる可能性 もある。新たに形成された集団が小さい場合、その創始者たちは、はるか未来にわたって集団の遺伝的構成に強い影響を与える可能性がある。 よく知られた例としては、1744年にペンシルベニア州に移住したアーミッシュ族がある。この新コロニーの2人のメンバーは、エリス・ヴァン・クレヴェル ド症候群の劣性対立遺伝子を共有していた。このコロニーのメンバーとその子孫は、宗教的に孤立し、比較的閉鎖的な傾向にある。何世代にもわたる近親交配の 結果、エリス・ヴァン・クレヴェルド症候群は、現在ではアーミッシュ族の間で一般人口よりもはるかに多く見られるようになっている。 元の集団とコロニーの遺伝子頻度の違いも、多くの世代を経るうちに2つの集団が大きく分岐するきっかけとなる可能性がある。この違い、すなわち遺伝的距離 が大きくなると、2つの分離した集団は遺伝的にも表現型においても明確に異なるようになる可能性がある。ただし、この分岐には遺伝的浮動だけでなく、自然 淘汰、遺伝子流動、突然変異も影響している。この集団の遺伝子頻度の比較的急速な変化の可能性により、ほとんどの科学者は、創始者効果(および拡張して遺 伝的浮動)を新種の進化における重要な推進力であると考えるようになった。ソール・ライトは、自身の「種の分化の平衡説」において、無作為な浮動と新たに 孤立した小規模な集団にこのような重要性を初めて関連付けた人物である。[46]ライトに続き、エルンスト・マイヤーは、創始者効果による遺伝的多様性の 減少と小規模な 創始者効果に続く遺伝的多様性の低下と個体数の少なさは、新種の進化にとって決定的に重要であることを示す説得力のあるモデルを数多く生み出した。 [47] しかし、この見解は今日ではあまり支持されていない。なぜなら、この仮説は実験的研究によって繰り返し検証されてきたが、結果はせいぜい曖昧なものでしか なかったからだ。[48] |
| History Further information: Modern synthesis (20th century) The role of random chance in evolution was first outlined by Arend L. Hagedoorn and Anna Cornelia Hagedoorn-Vorstheuvel La Brand in 1921.[49] They highlighted that random survival plays a key role in the loss of variation from populations. Fisher (1922) responded to this with the first, albeit marginally incorrect, mathematical treatment of the "Hagedoorn effect".[50] Notably, he expected that many natural populations were too large (an N ~10,000) for the effects of drift to be substantial and thought drift would have an insignificant effect on the evolutionary process. The corrected mathematical treatment and term "genetic drift" was later coined by a founder of population genetics, Sewall Wright. His first use of the term "drift" was in 1929,[51] though at the time he was using it in the sense of a directed process of change, or natural selection. Random drift by means of sampling error came to be known as the "Sewall–Wright effect," though he was never entirely comfortable to see his name given to it. Wright referred to all changes in allele frequency as either "steady drift" (e.g., selection) or "random drift" (e.g., sampling error).[52] "Drift" came to be adopted as a technical term in the stochastic sense exclusively.[53] Today it is usually defined still more narrowly, in terms of sampling error,[54] although this narrow definition is not universal.[55][56] Wright wrote that the "restriction of "random drift" or even "drift" to only one component, the effects of accidents of sampling, tends to lead to confusion".[52] Sewall Wright considered the process of random genetic drift by means of sampling error equivalent to that by means of inbreeding, but later work has shown them to be distinct.[57] In the early days of the modern evolutionary synthesis, scientists were beginning to blend the new science of population genetics with Charles Darwin's theory of natural selection. Within this framework, Wright focused on the effects of inbreeding on small relatively isolated populations. He introduced the concept of an adaptive landscape in which phenomena such as cross breeding and genetic drift in small populations could push them away from adaptive peaks, which in turn allow natural selection to push them towards new adaptive peaks.[58] Wright thought smaller populations were more suited for natural selection because "inbreeding was sufficiently intense to create new interaction systems through random drift but not intense enough to cause random nonadaptive fixation of genes".[59] Wright's views on the role of genetic drift in the evolutionary scheme were controversial almost from the very beginning. One of the most vociferous and influential critics was colleague Ronald Fisher. Fisher conceded genetic drift played some role in evolution, but an insignificant one. Fisher has been accused of misunderstanding Wright's views because in his criticisms Fisher seemed to argue Wright had rejected selection almost entirely. To Fisher, viewing the process of evolution as a long, steady, adaptive progression was the only way to explain the ever-increasing complexity from simpler forms. But the debates have continued between the "gradualists" and those who lean more toward the Wright model of evolution where selection and drift together play an important role.[60] In 1968, Motoo Kimura rekindled the debate with his neutral theory of molecular evolution, which claims that most of the genetic changes are caused by genetic drift acting on neutral mutations.[5][6] The role of genetic drift by means of sampling error in evolution has been criticized by John H. Gillespie[61] and William B. Provine, who argue that selection on linked sites is a more important stochastic force. |
歴史 詳細情報:現代の総合(20世紀) 進化におけるランダムな偶然の役割は、1921年にArend L. HagedoornとAnna Cornelia Hagedoorn-Vorstheuvel La Brandによって初めて概説された。[49] 彼らは、個体群の変異の喪失においてランダムな生存が重要な役割を果たしていることを強調した。フィッシャー(1922年)は、この「ヘゲドルン効果」に 対して、限定的ではあるが数学的に初めて取り扱った。[50] 注目すべきは、彼は多くの自然集団が大きすぎる(N=10,000)ため、ドリフトの影響が顕著ではないと予想し、ドリフトは進化プロセスにほとんど影響 を与えないと考えていたことだ。後に、集団遺伝学の創始者であるソール・ライトが、修正された数学的処理と「遺伝的浮動」という用語を考案した。彼が「ド リフト」という用語を初めて使用したのは1929年のことだったが[51]、当時は変化の方向づけられたプロセス、すなわち自然淘汰という意味で使用して いた。サンプリングエラーによるランダムなドリフトは「Sewall-Wright効果」として知られるようになったが、彼は自分の名前が付けられること にまったく快く思っていなかった。ライトは対立遺伝子頻度のあらゆる変化を「定常ドリフト」(例:選択)または「ランダムドリフト」(例:サンプリングエ ラー)と呼んだ。[52] 「ドリフト」は、もっぱら確率的な意味での専門用語として採用されるようになった。[53] 今日では、通常、サンプリングエラーという観点から、さらに狭義に定義されるが、この狭義の定義は普遍的なものではない 。[55][56]ライトは、「『ランダムドリフト』、あるいは『ドリフト』をサンプリングの偶然による影響という1つの要素のみに限定することは、混乱 を招く傾向がある」と記している。[52] ソウォール・ライトは、サンプリングエラーによるランダムな遺伝的ドリフトのプロセスを、近交によるものと同等のものとみなしたが、その後の研究により、 両者は異なるものであることが示されている。[57] 現代の進化総合説の初期、科学者たちは、チャールズ・ダーウィンの自然淘汰説に、集団遺伝学という新しい科学を融合させ始めていた。この枠組みの中で、ラ イトは、比較的小規模で孤立した集団における近親交配の影響に注目した。彼は、小規模な集団における交雑や遺伝的浮動などの現象が、適応度ピークからそれ らを遠ざける可能性があるという適応地形の概念を導入した。これにより、自然淘汰がそれらを新たな適応度ピークに向かって押し進めることができる。ライト は、小規模な集団は自然淘汰により適していると考えた。なぜなら、「近親交配は、ランダムな浮動により新たな相互作用システムを生み出すには十分に激しい が、ランダムな非適応遺伝子の固定を引き起こすほど激しいものではない」からだ。 進化の過程における遺伝的浮動の役割に関するライトの見解は、ほぼ当初から論争の的であった。最も声高で影響力のある批判者の一人は、同僚のロナルド・ フィッシャーであった。フィッシャーは、遺伝的浮動が進化において何らかの役割を果たしていることは認めたが、それは取るに足らないものだと考えていた。 フィッシャーは、ライトの見解を誤解していると非難された。なぜなら、フィッシャーの批判は、ライトが淘汰をほぼ完全に否定しているかのように思われたか らである。フィッシャーにとって、進化の過程を長い、安定した、適応的な進歩と捉えることは、単純な形態から複雑さが増し続けることを説明する唯一の方法 であった。しかし、「漸進論者」と、淘汰とドリフトが共に重要な役割を果たすライトの進化モデルに傾倒する人々との間の論争は続いた。 1968年、木村資生は分子進化の中立理論を提唱し、この理論では、遺伝的変化のほとんどは中立突然変異に作用する遺伝的浮動によって引き起こされると主張している。 進化におけるサンプリングエラーによる遺伝的浮動の役割については、ジョン・H・ギレスピーとウィリアム・B・プロバインから批判されている。彼らは、関連部位における選択の方が、より重要な確率的な力であると主張している。 |
| Peripatric speciation Antigenic drift Coalescent theory Constructive neutral evolution Gene pool Meiotic drive |
周辺種分化 抗原ドリフト 合同説 建設的中立進化 遺伝子プール 減数分裂駆動 |
| https://en.wikipedia.org/wiki/Genetic_drift |
|
| Hey, Jody, 2005. "On the Number of New World Founders: A Population
Genetic Portrait of the Peopling of the Americas" in PLoS Biol 2005 May
24;3(6):e193 DOI: 10.1371/journal.pbio.0030193 The founding of New World populations by Asian peoples is the focus of considerable archaeological and genetic research, and there persist important questions on when and how these events occurred. Genetic data offer great potential for the study of human population history, but there are significant challenges in discerning distinct demographic processes. A new method for the study of diverging populations was applied to questions on the founding and history of Amerind-speaking Native American populations. The model permits estimation of founding population sizes, changes in population size, time of population formation, and gene flow. Analyses of data from nine loci are consistent with the general portrait that has emerged from archaeological and other kinds of evidence. The estimated effective size of the founding population for the New World is fewer than 80 individuals, approximately 1% of the effective size of the estimated ancestral Asian population. By adding a splitting parameter to population divergence models it becomes possible to develop detailed portraits of human demographic history. Analyses of Asian and New World data support a model of a recent founding of the New World by a population of quite small effective size. |
アジアの人々による新世界人口の創設は、考古学および遺伝学の研究の焦
点となっており、これらの出来事がいつ、どのようにして起こったのかという重要な疑問が依然として残っている。遺伝子データは、人類集団の歴史研究に大き
な可能性を提供するが、個別の人口動態プロセスを識別するには大きな課題がある。分岐した集団の研究に新しい手法が適用され、アメリカインディアン語族を
話すネイティブアメリカン集団の起源と歴史に関する疑問が解明された。このモデルでは、集団の起源の規模、集団規模の変化、集団形成の時期、遺伝子流動の
推定が可能である。9つの遺伝子座のデータ分析は、考古学的証拠やその他の証拠から得られた一般的な見解と一致している。推定される新世界開拓集団の実効
集団サイズは80人以下であり、推定されるアジアの祖先集団の実効集団サイズの約1%である。分岐パラメータを集団分岐モデルに追加することで、人類の人
口統計学的歴史の詳細な描写が可能になる。アジアと新世界のデータの分析は、実効集団サイズが非常に小さい集団による新世界の最近の開拓というモデルを裏
付けている。 |
リンク
文献
その他の情報
Copyleft, CC, Mitzub'ixi Quq Chi'j, 1996-2099
![]()