
第6章 ある科学論文の解説:「実験室における社会実践の民族誌学的研究」
池田光穂・七五三木聡※
前章(→5章:リンク先) では、佐藤宏道のアメリカ留学におけるメンターであったナイジェル・ドウ教授のところに、七五三木聡を派遣され、論文が作成される背景のドラマを佐藤じし んの記述によって再現した。そこでは、科学者たちが実験をおこない論文を書くためには、さまざまな研究のネットワークの存在、実験を支える研究資金、研究 支援体制があることが示唆された。このようにその背景にある豊富な社会的実践の豊富さと眼もくらむばかりの多様性や多元性に比べると、論文執筆の知的作業 などは、それらの総合的な知的実践のほんの一部しか形成していないことは明らかである。
にもかかわらず、それらの広大な社会実践に肉薄するためには、まず個々の研究者の知的な作業の結実物である論文が具体的に何を意味するのかについ て知らなければならない。そこで本章では、七五三木聡らによる当該論文「サイクリックAMP依存プロテインキナーゼの阻害作用はネコの視覚皮質における逆 眼球優位を防ぐことができない」(Shimegi S, Fischer QS, Yang Y, Sato H, Daw NW. Blockade of cyclic AMP-dependent protein kinase does not prevent the reverse ocular dominance shift in kitten visual cortex. J Neurophysiol. 2003 90(6):4027-32)[次の第7章に再録]を理解するために最低限必要な学問的解説をおこなう。
■脳の可塑性
脳の可塑性とは「発達段階の神経系が環境に応じて最適の処理システムを作り上げるために、よく使われるニューロンの回路の処理効率を高め、使われな い回路の効率を下げるという現象」のことであり「発達期の脳において顕著にこの性質が観察される」(「ある科学論文の誕生」より)。ニューロンの可塑性に ついてはカナダの心理学者ドナルド・ヘッブ(Donald O. Hebb, 1904-1985)が提唱したヘッブ法則が有力視されており、ある一定の刺激が来た場合にのみシナプスの可塑性ができる(協同性)、特定のシナプスにの み可塑性がみられ関係のないものに可塑性は生じない(入力特異性)および、弱い刺激でも他の刺激の助けをかりて可塑性を生じる(連合性)の3つの特性で脳 の可塑性を説明するものである。ヘッブの法則に修正を加えたのがコバリアンスで、これはシナプスの強化は、あるニューロンがシナプスを介して次のニューロ ンを刺激できるかどうかにかかっており、そのことの強化刺激つづくことと可塑性には関連性があるというものである。
■脳の可塑性が生じる過程
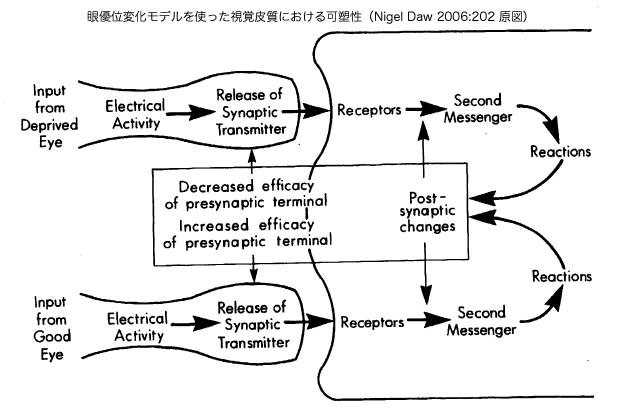
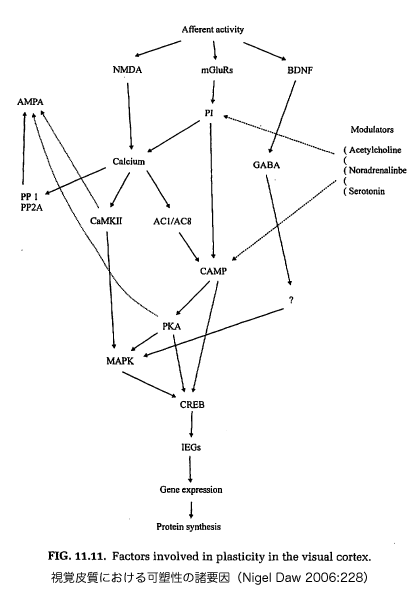
脳の可塑性を起こすためには、(興奮性)伝達物質のグルタミン酸と結合するNMDA受容体と呼ばれるグルタミン酸受容体の働きが重要とされている。 NMDAとは、N-methyl-D-aspartete の略号(アクロニム)であり、受容体の名前は、この受容体がNMDAと高い親和性(結合しやすい性質)示すことに由来する。NMDA受容体は、シナプス接 合部にあり、グルタミン酸と結合すると、興奮性の膜電位応答とともに、ニューロン内にカルシウムイオンが流入し、それが契機となって細胞内の酵素系——プ ロテインキナーゼA(Protain kinase A, PKA)もそのひとつ——が活性化され、化学反応を引き起こす。
■グルタミン酸受容体
グルタミン酸受容体は、その構造や性質からイオンチャンネル型受容体と代謝型受容体に分類されている。イオンチャンネル型受容体は、グルタミン酸と の結合によりその構造が変化し、イオンの通り道であるイオンチャンネルが開口することで、膜電位応答を引き起こす。NMDA[型]受容体は、イオンチャン ネル型受容体の一つであり、主に、ナトリウムイオンやカリウムイオンを通過させる一方で、カルシウムイオンも通過させることが特徴である。イオンチャンネ ル型受容体の中には、NMDA受容体以外にも、AMPA(α-amino-3-hydroxy-5methyl-isoxazole-4- propionate)やカイニン酸に親和性を有するAMPA/カイニン酸[型]受容体がある。ナトリウムイオンやカリウムイオンを通過させるが、カルシ ウムイオンは通過させないことが特徴である。AMPA/カイニン酸は、それ以外の性質においてもNMDA受容体との違いがみられることから、イオンチャン ネル型受容体内において両者を区別するために、non-NMDA受容体とも呼ばれる。 代謝型受容体は、それ自体がイオンチャンネルを持たず、G--プロテインと呼ばれる別のタンパク質などを介してイオンチャンネルの開閉や酵素活性の変化な どを引き起こす。
(→文献:Tumoto,T., Neurosci. Res., 9:79-102, 1990.)
■OD(ocular dominance)眼優位性
ネコの大脳皮質視覚野のニューロンの多くは左右どちらの眼に光刺激を与えても反応する性質(両眼反応性)を示すが、どちらの眼からの入力に対してよ り強く応じるかは、ニューロンによって異なる。ニューロンによっては片方の眼への光刺激のみに応じるものもある。このようなニューロンの眼に対する反応選 択性を眼優位性(ocular dominance: OD)と呼ぶ。
■臨界期(critical period)
生後発達の比較的早期に、環境からの影響を強く受け、動物と環境の相互作用に基づいて脳の構造および機能の発達が促される時期があり、これを臨界期あるいは感受性期と呼ぶ。臨界期の発育環境が異常であると、正常な発達が損なわれることが多い。
臨界期の始まる時期や継続する長さは、それぞれの機能ごとに、また、種によっても違いがみられる。大脳皮質視覚野における典型的な可塑性現象ある眼優位性シフトは、ネコの場合、4‾6週齢であり、七五三木論文では、6週齢のネコが使われている。
■眼優位性シフト:
脳の可塑性を示す実例の一つ。正常な視覚体験をして成長したネコの大脳皮質視覚野では、多くのニューロンが両眼反応性を有している。生後1ヶ月の仔 ネコでも、同様な両眼反応性が観察されるが、この時期に片眼を閉じて飼育する——これを片眼(単眼)遮蔽飼育という——と、ほとんどのニューロンは、閉じ た方の眼(遮閉眼)への光刺激には反応しなくなり、開いていた方の眼(健常眼)にのみ反応するようになる。このような眼に対する反応選択性の変化を眼優位 性シフト(ocular dominance shift: OD shift)と呼ぶ。このようなシフトは、臨界期中に、遮閉眼への正常な視覚入力を開始することで、両眼反応性が回復するが、臨界期の後に再開しても、遮 蔽眼に対する反応はほとんど回復しない。
■PKA(プロテインキナーゼ A)の阻害剤としてのRp-8-Cl-cAMPS
PKA(プロテインキナーゼ A)の阻害剤としてのRp-8-Cl-cAMPS をネコの皮質視覚野に慢性的に投与し続ける(chronic infusion)と、正常型眼優位性シフトが起こらなくなる。このことから、正常型眼優位性シフトの分子メカニズムとしてPKAが関与していることがわ かる。しかし、PKAがどのような経路で活性化されるのか、また、どのようにして眼優位性のシフトに関与し得るのかについては、現在でも不明のままであ る。
■2種類の眼優位性シフト
臨界期にある動物の大脳皮質視覚野ニューロンの眼優位性は、単眼遮蔽(monocular deprivation:MD)のような人為的な操作によって変化する。これを眼優位性シフトと呼ぶが、その変化の方向性は必ずしも一つではない。通常、 視覚遮蔽された眼(遮蔽眼)に対する反応性が失われ、ほとんどのニューロンが健常眼に対してのみに反応するようになる。これに対して、大脳皮質視覚野を不 活性化した状況下で単眼遮蔽を行うと、健常眼よりも遮蔽眼により強く反応するニューロンが増加する。大脳皮質の不活性化とは、皮質のニューロンが強い興奮 性入力を受けてもスパイク応答を生じないような状況にすることであり、一つの方法として、抑制性伝達物質GABAの受容体アゴニスト(作動薬)ムシモール を視覚皮質に持続注入することで皮質のニューロンが抑制され、不活性化の状況が得られる。この大脳皮質視覚野の不活性化と単眼遮蔽の組み合わせは、一見不 自然とも思えるような方向へのシフト、すなわち、遮蔽眼への選択性が高まるようなシフトを引き起こす。これを逆方向性眼優位性シフト(reverse ocular dominance shift)と呼び、これとの比較の意味で、前者のシフトを正常型眼優位性シフト(normal ocular dominance shift)と呼ぶ。
逆方向性眼優位性シフト現象は、大脳皮質視覚野ニューロンの眼優位性の変化が、単純にどちらの眼からの視覚入力が強いか弱いかによって決定されるも のではないことを示している。現在、心理学者のドナルド・ヘッブにより提唱されたヘッブの法則(ヘッブ則)に修正を加えたコバリアンス則と呼ばれる仮説 が、これらの現象を統一的に説明するものとして有力視されている。これによれば、ニューロンが発火した時、その時に入力を与えていたシナプスが強化され、 逆に、発火していない時に入力を与えていたシナプスは退縮していく。言い換えれば、あるシナプスが強化されるか退縮するかは、そのシナプスからの入力に よって次のニューロンを発火させられるかどうかということになる。発火させられれば、その情報は、さらに次のニューロンへと伝達されることを意味し、その シナプスは情報処理の一部を担ったことになり、そのシナプスの強化は情報処理の効率化へとつながる。逆に、シナプスから入力が与えられても、次のニューロ ンが発火しない状態が続く場合、その入力はニューロンにとって不必要ものであり、その入力を減弱させることで情報処理の最適化を図ると考えられている。正 常型眼優位性シフトは前者のケースとして、また、逆方向性眼優位性シフトは後者のケースとして説明され得る。
■NMDA受容体の特徴
NMDA受容体のイオンチャネル機能は、ニューロンが興奮していない状態では、細胞外からのマグネシウムイオンによってブロックされている。チャ ンネル機能が発現するためには、グルタミン酸との結合とともに、AMPA受容体などを介した細胞膜の興奮によってマグネシウムによるブロックが解除される 必要がある。そのため、ニューロンに興奮性入力が与えられても弱い興奮ではNMDA受容体が機能せず、さらに強い入力が与えられ、より強い興奮が生じた時 のみに働くことになる。NMDA受容体が働けば、さらに大きな興奮がニューロンに生じ、それとともに、種々の細胞機能を調節するトリッガーとしての働きを もつカルシウムが流入する。これは、ニューロンの興奮の程度がNMDA受容体を活性化できるレベルより上か下かによって、ニューロンに与える影響が大きく 異なることを意味する。このようなNMDA受容体の働きは、強い入力を与えるシナプスを選択的に強化するための仕組みとしてシナプス可塑性に関与する可能 性が指摘されている。NMDA受容体は眼優位性シフトの発現にも関与することが示唆されており、臨界期の動物の視覚野ではNMDA受容体の機能が高まるこ と、また、片眼遮蔽による正常型眼優位性シフトは、大脳皮質視覚野にNMDA受容体の拮抗薬を持続注入することで阻害されることが知られている。
■MD(monocular deprivation)単眼遮蔽・片眼遮蔽
片方のまぶたを縫合したり、眼帯などにより遮光することで、片方の眼への視覚入力を遮蔽すること。臨界期に単眼遮蔽を行うと大脳皮質視覚野のニューロンに眼優位性シフトが起きるが、臨界期以降であれば、単眼遮蔽してもそのような変化は起こらない。
■ムシモール(muscimol)
ムシモール(化学式:C4H6N2O2)は、イボテン酸が脱炭酸したものであり、ベニテングタケなどに含まれる毒成分であると言われる 。抑制性の神経伝達物質の一つであるγアミノ酪酸(GABA)と構造が類似し、GABAa受容体に結合することで、ニューロンの興奮性を低下させ、スパイ ク応答を生じにくくさせる(不活性化)。脳が実際に伝達物質として利用するGABAには分解酵素がシナプス部分に存在するので、放出されたGABAは速や かに分解される。これに対して、ムシモールは、そのような分解酵素が効かないため残存時間が長く、長時間に渡って、ニューロンを不活性化するのに利用され ることが多い。
■ラトゥール『科学が作られているとき』(1999[1987])における〈既製の科学〉と〈作成過程の科学〉という2つの面をもつヤヌス像
既製の科学 作成過程の科学
・ただ事実をまっすぐに得よ!(p.13)
・最も効率的な機械を手にいれよ(p.16)。
・機械がひとたび作動すると、人々は確信するようになる(p.19)。
・事物が真であれば、それらはもちこたえる(p.21)。
・科学は多数意見によって曲げられない(p.53)。
・自然は論争を決着させる原因である(p.170)。
・社会は論争を決着させる原因である。(p.248)
・科学と技術はプロジェクトを実行可能にする原因である(p.302)。
___________________
・役に立たない事実はすべて取り除け。
・効率性とは何であるべきかに基づいて決めよ。
・すべての関係者が確信すると、機械は作動するようになる。
・事実がもちこたえれば、それらは真になり始める。
・どうしたら多数意見よりも強くなるだろうか。
・自然は決着の結果として存在することになろう。
・社会の安定状態は論争の決着の結果として存在することになろう。
・プロジェクトが実行されたとき科学と技術はその背後の推進力のように見える。
■事実A
正常型眼優位性シフトは、PKAの阻害剤であるRp-8-Cl-cAMPSによって完全に阻害される。では逆方向性眼優位性シフトは、Rp-8- Cl-cAMPS によって阻害されるのか? 逆方向性眼優位シフトは、仔ネコの視覚皮質に、GABA受容体の作動薬であるムシモールを持続注入して皮質を不活性化した状態 で、単眼遮蔽を行うことで得られるので、この仮説を証明するためには、この条件に加えてRp-8-Cl-cAMPSをムシモールと共に視覚皮質へ慢性的に 流し続けるという実験を行う必要がある。七五三木論文はこれを行ったものである。では、その結果はいかなるものであったのか?逆方向性眼優位シフトは邪魔 されなかった。つまりRp-8-Cl-cAMPSの存在下でPKAが活性化しない状況下にあっても起こってしまったのである。これは、逆方向性眼優位性シ フトにPKAが関与していないことを強く示唆している。それとともに、臨界期に観察される2種類の眼優位性シフトの分子メカニズムは同じではなく、正常型 眼優位性シフトはPKAを介した細胞内情報伝達系を介して、一方、逆方向性眼優位性シフトはそれ以外の経路でシフト現象を起こすことが示唆される。
■事実B
PKAの作用を阻害するRp-8-Cl-cAMPSの存在の有無にもかかわらず、逆方向性眼優位シフトは起こる。大脳皮質の層構造との関係の中で、 シフトの程度を比較すると、大脳皮質の入力層(視床外側膝状体から直接入力を受ける層)であるIV層において最も強い眼優位が観察された。ネコの場合、右 眼と左眼からの視覚情報は、大脳皮質視覚野のIV層で初めて一つのニューロンに収束し、それによりそれらのニューロンは両眼反応性を獲得する。しかし、そ れ以前の段階では、左右それぞれの眼からの情報は、眼優位性が異なるニューロン群によって別々に処理・伝達されている。IV層でシフトが大きいということ は、視床外側膝状体から大脳皮質視覚野ニューロンへと投射する線維の形成するシナプス(thalamocortical synapses)の変化が眼優位性シフトの神経学的基礎になっている可能性を示唆する。一方、大脳皮質視覚野内では、IV層→II・III層→V層 →VI層のような情報の流れがあり、その過程で、情報の収束が起こることから、遮蔽眼へ強い選択性を示すニューロンに加え、ある程度健常眼にも応答する ニューロンらがIV層以降の段階で収束することによって、遮蔽眼への反応選択性が減弱するものと思われる。
■著者たちの推論
著者たちの結論はこうである。(1)逆方向性眼優位シフトが生じる過程に、PKAの活性化は必要でない、(2)単眼遮蔽によって生じる眼優位性可塑 性の分子メカニズム、すなわち、細胞内情報伝達機構は一つではなく複数ある。少なくとも、大脳皮質視覚野が正常な状態で単眼遮蔽することで起こる正常型眼 優位シフトはPKAの活性化を必要とし、これを必要としない逆方向性眼優位シフトとは分子メカニズムが異なると考えられる。
□ポストスクリプト
七五三木の論文を理解することとはどうゆうことであろうか?
——七五三木という科学者の個人や彼の研究史を理解すればわかる?:いいえ。
——論文に引用されているすべての文献を読破すればわかる?:いいえ。
——視覚の神経生理の教科書やレビュー(e.g. Daw 2006)を読めばわかる?:はい、しかしながら関連文献や生理学に関する知識も必要になるであろう。
七五三木の論文を理解することは、論文に記載されたこれまでの科学的知見を集約するだけでなく論文の著者たちが、どのような新知見をこの論文に込 めようとしたのか、またこの研究パラダイムにおいて焦点化されている「パズル」とは何かということを的確に知ることである。七五三木の論文を短期間で理解 するためには、オッカムの剃刀のごとく問題にもとづく学習(problem-based learning)のような焦点化された学習が必要である。しかし、それだけでは視覚の神経生理学の全体像はおぼろげにしか見えてこない。結局のところ は、その研究の全体像を理解するためには、広範な文献渉猟と研究者どうしの絶え間のない対話に参加するしかないということになる。
■Signal transduction[資料]
In biology, signal transduction refers to any process by which a cell converts one kind of signal or stimulus into another. Most processes of signal transduction involve ordered sequences of biochemical reactions inside the cell, which are carried out by enzymes, activated by second messengers, resulting in a signal transduction pathway. Such processes are usually rapid, lasting on the order of milliseconds in the case of ion flux, or minutes for the activation of protein- and lipid-mediated kinase cascades, but some can take hours, and even days (as is the case with gene expression), to complete. The number of proteins and other molecules participating in the events involving signal transduction increases as the process emanates from the initial stimulus, resulting in a "signal cascade," beginning with a relatively small stimulus that elicits a large response. This is referred to as amplification of the signal. source: http://en.wikipedia.org/wiki/Image:Signal_transduction_v1.png
文献
- Daw, Nigel W., 2006. Visual Development. 2nd ed., New York: Springer.
- ラトゥール、B., 1999『科学が作られているとき:人類学的考察』川崎勝・高田紀代志訳、産業図書。
※本章は池田光穂が執筆し七五三木が、学術的な観点から校閲した。もし誤りがあれば、それは七五三木のせいではなく、その文責は池田光穂にある。
◆「実験室における社会実践の民族誌学的研究」(目次)
Copyleft, CC, Mitzub'ixi Quq Chi'j, 1996-2099