Neural Correlates of Consciousness, NCC
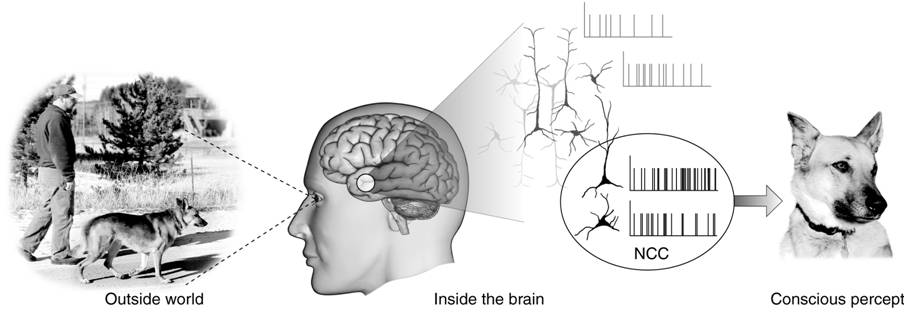
意識の神経相関
Neural Correlates of Consciousness, NCC
解説:池田光穂
「特定の意識経験」に対応する神経活動や パターンのことをいう。
しかしながら、この想定や存在論的根拠に は種々の議論があり決着がついているわけではない。
| The neural
correlates of consciousness (NCC) are the minimal set of neuronal
events and mechanisms sufficient for the occurrence of the mental
states to which they are related.[2] Neuroscientists use empirical
approaches to discover neural correlates of subjective phenomena; that
is, neural changes which necessarily and regularly correlate with a
specific experience.[3][4] The set should be minimal because, under the
materialist assumption that the brain is sufficient to give rise to any
given conscious experience, the question is which of its components are
necessary to produce it. |
意識の神経相関(NCC)とは、関連する精神状態の発生に十分な最小限
の神経細胞の事象およびメカニズムである。神経科学者は、主観的現象の神経相関を発見するために経験的アプローチを用いる。つまり、特定の経験と必然的か
つ規則的に相関する神経変化である。このセットは最小限でなければならない。なぜなら、脳が意識経験を生み出すのに十分であるという唯物論的仮定のもとで
は、その構成要素のうちどれが意識経験を生み出すのに必要であるかが問題となるからだ。 |
The neuronal correlates of consciousness (NCC) constitute the smallest set of neural events and structures sufficient for a given conscious percept or explicit memory. This case involves synchronized action potentials in neocortical pyramidal neurons.[1] |
意識の神経相関(NCC)は、与えられた意識的知覚や明示的記憶に十分な、神経事象と神経構造の最小セットを構成する。この場合、大脳新皮質の錐体ニュー ロンにおける活動電位が同期している[1]。 |
| Neurobiological approach to
consciousness A science of consciousness must explain the exact relationship between subjective mental states and brain states, the nature of the relationship between the conscious mind and the electrochemical interactions in the body (mind–body problem). Progress in neuropsychology and neurophilosophy has come from focusing on the body rather than the mind. In this context the neuronal correlates of consciousness may be viewed as its causes, and consciousness may be thought of as a state-dependent property of an undefined complex, adaptive, and highly interconnected biological system.[5] Discovering and characterizing neural correlates does not offer a causal theory of consciousness that can explain how particular systems experience anything, the so-called hard problem of consciousness,[6] but understanding the NCC may be a step toward a causal theory. Most neurobiologists propose that the variables giving rise to consciousness are to be found at the neuronal level, governed by classical physics. There are theories proposed of quantum consciousness based on quantum mechanics.[7] There is an apparent redundancy and parallelism in neural networks so, while activity in one group of neurons may correlate with a percept in one case, a different population may mediate a related percept if the former population is lost or inactivated. It may be that every phenomenal, subjective state has a neural correlate. Where the NCC can be induced artificially, the subject will experience the associated percept, while perturbing or inactivating the region of correlation for a specific percept will affect the percept or cause it to disappear, giving a cause-effect relationship from the neural region to the nature of the percept. Proposals that have been advanced over the years include: what characterizes the NCC? What are the commonalities between the NCC for seeing and for hearing? Will the NCC involve all the pyramidal neurons in the cortex at any given point in time? Or only a subset of long-range projection cells in the frontal lobes that project to the sensory cortices in the back? Neurons that fire in a rhythmic manner? Neurons that fire in a synchronous manner?[8] The growing ability of neuroscientists to manipulate neurons using methods from molecular biology in combination with optical tools (e.g., Adamantidis et al. 2007) depends on the simultaneous development of appropriate behavioral assays and model organisms amenable to large-scale genomic analysis and manipulation. It is the combination of fine-grained neuronal analysis in animals with increasingly more sensitive psychophysical and brain imaging techniques in humans, complemented by the development of a robust theoretical predictive framework, that will hopefully lead to a rational understanding of consciousness, one of the central mysteries of life. |
意識に対する神経生物学的なアプローチ 意識の科学は、主観的な精神状態と脳の状態の正確な関係、意識と体内の電気化学的相互作用の関係性(心身問題)を説明しなければならない。神経心理学と神 経哲学の進歩は、精神よりも身体に焦点を当てることでもたらされた。この文脈において、意識の神経相関は、その原因と見なされる可能性があり、意識は、未 定義の複雑かつ適応性があり、高度に相互接続された生物学的システムの、状態依存的な特性であると考えられる可能性がある。 神経相関を発見し、特徴づけることは、特定のシステムがどのようにして何かを経験するのかを説明できる意識の因果理論、いわゆる意識の難問[6]を提供す るものではないが、NCCを理解することは因果理論への一歩となる可能性がある。ほとんどの神経生物学者は、意識を生み出す変数は古典物理学によって支配 される神経細胞レベルにあると提唱している。量子力学に基づく量子意識の理論も提唱されている。 神経ネットワークには明らかな冗長性と並列性があるため、あるニューロン群の活動が知覚と相関する一方で、前者のニューロン群が失われたり不活性化したり した場合、別の集団が関連する知覚を媒介することがある。あらゆる現象的、主観的な状態には、神経相関があるのかもしれない。NCCが人工的に誘導される 場合、被験者は関連する知覚を経験する。一方、特定の知覚に関連する領域を撹乱または不活性化すると、その知覚に影響を与えたり、知覚を消失させたりす る。これにより、神経領域から知覚の性質に至る因果関係が明らかになる。 長年にわたって提案されてきたものには、次のようなものがある。NCCの特徴とは何か?視覚と聴覚におけるNCCの共通点とは何か?NCCは、ある特定の 時点において、大脳皮質にあるすべての錐体ニューロンを含むのか?それとも、後部の感覚皮質に投射する前頭葉にある長距離投射細胞のサブセットだけなの か?リズムを刻むように発火するニューロン?同期して発火するニューロン?[8] 分子生物学的手法と光学ツール(例えば、Adamantidis et al. 2007)を組み合わせた神経科学者による神経細胞の操作能力の向上は、大規模なゲノム解析と操作に適した適切な行動アッセイとモデル生物の同時開発に依 存している。動物におけるきめ細かな神経細胞分析と、人間におけるますます高感度な心理物理学および脳画像化技術の組み合わせ、そして強固な理論予測フ レームワークの開発によって補完されることが、生命の中心的な謎のひとつである意識の合理的な理解につながるだろう。 |
| Level of arousal and content of
consciousness There are two common but distinct dimensions of the term consciousness,[9] one involving arousal and states of consciousness and the other involving content of consciousness and conscious states. To be conscious of anything the brain must be in a relatively high state of arousal (sometimes called vigilance), whether in wakefulness or REM sleep, vividly experienced in dreams although usually not remembered. Brain arousal level fluctuates in a circadian rhythm but may be influenced by lack of sleep, drugs and alcohol, physical exertion, etc. Arousal can be measured behaviorally by the signal amplitude that triggers some criterion reaction (for instance, the sound level necessary to evoke an eye movement or a head turn toward the sound source). Clinicians use scoring systems such as the Glasgow Coma Scale to assess the level of arousal in patients. High arousal states are associated with conscious states that have specific content, seeing, hearing, remembering, planning or fantasizing about something. Different levels or states of consciousness are associated with different kinds of conscious experiences. The "awake" state is quite different from the "dreaming" state (for instance, the latter has little or no self-reflection) and from the state of deep sleep. In all three cases the basic physiology of the brain is affected, as it also is in altered states of consciousness, for instance after taking drugs or during meditation when conscious perception and insight may be enhanced compared to the normal waking state. Clinicians talk about impaired states of consciousness as in "the comatose state", "the persistent vegetative state" (PVS), and "the minimally conscious state" (MCS). Here, "state" refers to different "amounts" of external/physical consciousness, from a total absence in coma, persistent vegetative state and general anesthesia, to a fluctuating and limited form of conscious sensation in a minimally conscious state such as sleep walking or during a complex partial epileptic seizure.[10] The repertoire of conscious states or experiences accessible to a patient in a minimally conscious state is comparatively limited. In brain death there is no arousal, but it is unknown whether the subjectivity of experience has been interrupted, rather than its observable link with the organism. Functional neuroimaging have shown that parts of the cortex are still active in vegetative patients that are presumed to be unconscious;[11] however, these areas appear to be functionally disconnected from associative cortical areas whose activity is needed for awareness. The potential richness of conscious experience appears to increase from deep sleep to drowsiness to full wakefulness, as might be quantified using notions from complexity theory that incorporate both the dimensionality as well as the granularity of conscious experience to give an integrated-information-theoretical account of consciousness.[12] As behavioral arousal increases so does the range and complexity of possible behavior. Yet in REM sleep there is a characteristic atonia, low motor arousal and the person is difficult to wake up, but there is still high metabolic and electric brain activity and vivid perception. Many nuclei with distinct chemical signatures in the thalamus, midbrain and pons must function for a subject to be in a sufficient state of brain arousal to experience anything at all. These nuclei therefore belong to the enabling factors for consciousness. Conversely, it is likely that the specific content of any particular conscious sensation is mediated by particular neurons in the cortex and their associated satellite structures, including the amygdala, thalamus, claustrum and the basal ganglia. |
覚醒レベルと意識の内容 意識という用語には、2つの共通するが異なる次元がある。[9] 1つは覚醒と意識の状態を伴うもので、もう1つは意識の内容と意識の状態を伴うものである。 何かを意識するためには、脳は覚醒(警戒とも呼ばれる)状態にある必要がある。これは、覚醒時でもレム睡眠時でも同様である。夢の中で鮮明に経験するが、 通常は記憶されない。脳の覚醒レベルは概日リズムで変動するが、睡眠不足、薬物、アルコール、身体的労作などによって影響を受ける可能性がある。覚醒は、 何らかの基準となる反応(例えば、眼球運動を引き起こすのに必要な音の大きさや、音源に向かって頭を回す)を引き起こす信号振幅によって行動的に測定する ことができる。臨床医は、グラスゴー昏睡尺度などのスコアリングシステムを使用して、患者の覚醒レベルを評価する。 高い覚醒状態は、特定の内容を持つ意識状態、すなわち、何かを見たり、聞いたり、記憶したり、計画したり、空想したりする意識状態と関連している。異なる 意識レベルや意識状態は、異なる種類の意識体験と関連している。「覚醒」状態は、「夢見」状態(例えば、後者は自己反省をほとんど、あるいはまったく伴わ ない)や深い睡眠状態とはまったく異なる。いずれの場合も、脳の生理学的な基本機能に影響が及ぶが、それは意識変容状態、例えば薬物の摂取後や瞑想中の意 識状態が、通常の覚醒状態と比較して、意識的な知覚や洞察力が強化される場合にも見られる。 臨床医は、意識障害状態を「昏睡状態」、「持続性植物状態」(PVS)、「最小意識状態」(MCS)などと表現する。ここでいう「状態」とは、昏睡状態、 持続性植物状態、全身麻酔状態のように意識が完全にない状態から、夢遊病や複雑部分てんかん発作中の意識のように、意識の感覚が変動し限定的な状態まで、 外部/物理的な意識の「量」の違いを指す。 最小意識状態にある患者が経験できる意識状態や体験のレパートリーは比較的限られている。脳死状態では覚醒はないが、経験の主観性が遮断されているのか、 あるいは生物としての観察可能なつながりが遮断されているのかは不明である。機能的ニューロイメージングでは、無意識であると推定される植物状態の患者で も大脳皮質の部分が依然として活動していることが示されているが[11]、これらの領域は意識に必要な活動を行う連合皮質領域とは機能的に遮断されている ようである。 意識体験の潜在的な豊かさは、複雑系理論の概念を用いて定量化できる可能性があるように思われる。この理論は、意識体験の次元性と粒度を統合情報理論的に 説明するために、意識体験の両方を組み込んでいる。[12] 行動の覚醒が高まるにつれ、可能な行動の範囲と複雑性も高まる。しかし、レム睡眠には特徴的な弛緩、低運動覚醒があり、人は目覚めにくくなるが、代謝や脳 の電気的活動は活発で、知覚も鮮明である。 視床、中脳、橋には、それぞれ異なる化学的特徴を持つ多くの神経核があり、被験者が何らかの経験をするのに十分な脳の覚醒状態にあるためには、それらの神 経核が機能している必要がある。したがって、これらの神経核は意識を生み出す要因に属する。逆に言えば、特定の意識的な感覚の具体的な内容は、大脳皮質 や、扁桃体、視床、クラウストラム、大脳基底核などの関連衛星構造にある特定のニューロンによって媒介されている可能性が高い。 |
| Neuronal basis of perception The possibility of precisely manipulating visual percepts in time and space has made vision a preferred modality in the quest for the NCC. Psychologists have perfected a number of techniques – masking, binocular rivalry, continuous flash suppression, motion induced blindness, change blindness, inattentional blindness – in which the seemingly simple and unambiguous relationship between a physical stimulus in the world and its associated percept in the privacy of the subject's mind is disrupted.[13] In particular a stimulus can be perceptually suppressed for seconds or even minutes at a time: the image is projected into one of the observer's eyes but is invisible, not seen. In this manner the neural mechanisms that respond to the subjective percept rather than the physical stimulus can be isolated, permitting visual consciousness to be tracked in the brain. In a perceptual illusion, the physical stimulus remains fixed while the percept fluctuates. The best known example is the Necker cube whose 12 lines can be perceived in one of two different ways in depth. 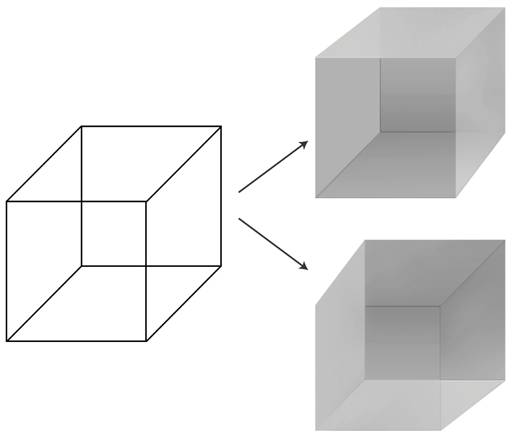 The Necker Cube: The left line drawing can be perceived in one of two distinct depth configurations shown on the right. Without any other cue, the visual system flips back and forth between these two interpretations.[14] A perceptual illusion that can be precisely controlled is binocular rivalry. Here, a small image, e.g., a horizontal grating, is presented to the left eye, and another image, e.g., a vertical grating, is shown to the corresponding location in the right eye. In spite of the constant visual stimulus, observers consciously see the horizontal grating alternate every few seconds with the vertical one. The brain does not allow for the simultaneous perception of both images. Logothetis and colleagues[15] recorded a variety of visual cortical areas in awake macaque monkeys performing a binocular rivalry task. Macaque monkeys can be trained to report whether they see the left or the right image. The distribution of the switching times and the way in which changing the contrast in one eye affects these leaves little doubt that monkeys and humans experience the same basic phenomenon. In the primary visual cortex (V1) only a small fraction of cells weakly modulated their response as a function of the percept of the monkey while most cells responded to one or the other retinal stimulus with little regard to what the animal perceived at the time. But in a high-level cortical area such as the inferior temporal cortex along the ventral stream almost all neurons responded only to the perceptually dominant stimulus, so that a "face" cell only fired when the animal indicated that it saw the face and not the pattern presented to the other eye. This implies that NCC involve neurons active in the inferior temporal cortex: it is likely that specific reciprocal actions of neurons in the inferior temporal and parts of the prefrontal cortex are necessary. A number of fMRI experiments that have exploited binocular rivalry and related illusions to identify the hemodynamic activity underlying visual consciousness in humans demonstrate quite conclusively that activity in the upper stages of the ventral pathway (e.g., the fusiform face area and the parahippocampal place area) as well as in early regions, including V1 and the lateral geniculate nucleus (LGN), follow the percept and not the retinal stimulus.[16] Further, a number of fMRI[17][18] and DTI experiments[19] suggest V1 is necessary but not sufficient for visual consciousness.[20] In a related perceptual phenomenon, flash suppression, the percept associated with an image projected into one eye is suppressed by flashing another image into the other eye while the original image remains. Its methodological advantage over binocular rivalry is that the timing of the perceptual transition is determined by an external trigger rather than by an internal event. The majority of cells in the inferior temporal cortex and the superior temporal sulcus of monkeys trained to report their percept during flash suppression follow the animal's percept: when the cell's preferred stimulus is perceived, the cell responds. If the picture is still present on the retina but is perceptually suppressed, the cell falls silent, even though primary visual cortex neurons fire.[21][22] Single-neuron recordings in the medial temporal lobe of epilepsy patients during flash suppression likewise demonstrate abolishment of response when the preferred stimulus is present but perceptually masked.[23] |
知覚の神経学的基礎 視覚知覚を時間的・空間的に正確に操作できる可能性があるため、視覚はNCCの探求において最も好ましい形態となっている。心理学者は、マスキング、両眼 競合、連続フラッシュ抑制、運動誘導性失明、変化盲、不注意盲など、いくつかの技術を完成させてきた。これらの技術では、世界の物理的刺激と、それに関連 する知覚との間に見られる一見単純かつ明確な関係が 被験者の心の中で混乱する。[13] 特に、刺激は一度に数秒間、あるいは数分間知覚的に抑制されることがある。つまり、画像は観察者の片方の目に投影されるが、見えないため、見えないのであ る。このようにして、物理的な刺激ではなく、主観的な知覚に反応する神経メカニズムを分離することができ、視覚的な意識を脳内で追跡することが可能にな る。知覚の錯覚では、物理的な刺激は固定されたままであるが、知覚は変動する。最もよく知られている例は、ネッカーの立方体(Necker cube)で、12本の線は奥行きにおいて2つの異なる方法で知覚される。 ネッカーの立方体:左の線画は、右に示す2つの異なる奥行き構成のうちの1つとして知覚される。他の手がかりがなければ、視覚システムはこれらの2つの解 釈の間を行ったり来たりする。[14] 正確に制御できる知覚の錯覚は、両眼競合である。ここでは、例えば水平の縞模様のような小さな画像が左目に提示され、別の画像、例えば垂直の縞模様が右目 の対応する位置に表示される。視覚刺激は常に同じであるにもかかわらず、観察者は水平の縞模様が数秒ごとに垂直の縞模様と交互に現れるように見える。脳は 両方の画像を同時に知覚することを許さない。 Logothetis氏とその同僚[15]は、両眼競合課題を行っている覚醒状態のマカクザルにおいて、さまざまな視覚皮質領域を記録した。マカクザル は、左または右の画像を見ているかを報告するように訓練することができる。切り替えにかかる時間の分布と、片方の目のコントラストを変えることでそれがど のように影響されるかを見ると、サルと人間が同じ基本的な現象を経験していることは疑いようがない。一次視覚野(V1)では、ほとんどの細胞が動物の知覚 とはほとんど関係なく、どちらかの網膜刺激に反応する一方で、ごく一部の細胞だけが、サルの知覚の機能として反応を弱く変調した。しかし、腹側経路に沿っ た下側頭葉皮質のような高次皮質領域では、ほぼすべてのニューロンが知覚的に優勢な刺激のみに反応し、動物が顔を見ていることを示した場合にのみ「顔」細 胞が反応した。このことは、NCCには下側頭葉皮質で活発に活動するニューロンが関与していることを示唆している。下側頭葉と前頭前皮質の一部のニューロ ンが相互に特定の作用を及ぼすことが必要である可能性が高い。 両眼競合や関連する錯視を利用し、人間の視覚意識の根底にある血流動態活動を特定するfMRI実験の多くは、腹側経路の上位段階(例えば、顔領域や海馬傍 領域)や、 V1や外側膝状体(LGN)などの初期領域は、知覚に従うものであり、網膜刺激に従うものではないことが示されている。[16] さらに、fMRI[17][18]やDTI[19]を用いた多くの実験から、V1は視覚意識には必要であるが、十分ではないことが示唆されている。 [20] 関連する知覚現象であるフラッシュ抑制では、片方の目に画像を投影すると、もう片方の目に別の画像を投影することで、元の画像を残したまま、その知覚が抑 制される。両眼競合に対する方法論的な利点は、知覚の変化のタイミングが内部イベントではなく外部トリガーによって決定されることである。フラッシュ抑制 中に知覚を報告するように訓練されたサルの下側頭皮質と上側頭溝の大部分の細胞は、動物の知覚に従う。細胞が好む刺激が知覚されると、細胞は反応する。網 膜にはまだ画像が残っているが知覚的には抑制されている場合、一次視覚野のニューロンが発火しても、その細胞は反応しなくなる。[21][22] 閃光抑制中のてんかん患者の内側側頭葉における単一ニューロン記録でも、好みの刺激が存在するが知覚的にはマスクされている場合、反応がなくなることが示 されている。[23] |
| Global disorders of consciousness Given the absence of any accepted criterion of the minimal neuronal correlates necessary for consciousness, the distinction between a persistently vegetative patient who shows regular sleep-wave transitions and may be able to move or smile, and a minimally conscious patient who can communicate (on occasion) in a meaningful manner (for instance, by differential eye movements) and who shows some signs of consciousness, is often difficult. In global anesthesia the patient should not experience psychological trauma but the level of arousal should be compatible with clinical exigencies. 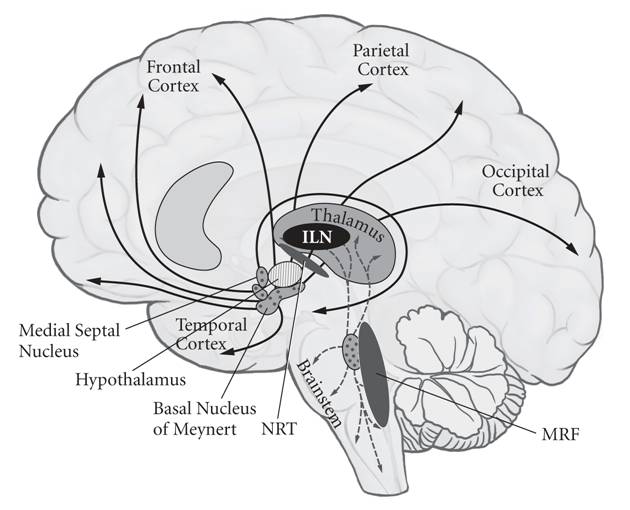 Midline structures in the brainstem and thalamus necessary to regulate the level of brain arousal. Small, bilateral lesions in many of these nuclei cause a global loss of consciousness.[24] Blood-oxygen-level-dependent fMRI have demonstrated normal patterns of brain activity in a patient in a vegetative state following a severe traumatic brain injury when asked to imagine playing tennis or visiting rooms in his/her house.[25] Differential brain imaging of patients with such global disturbances of consciousness (including akinetic mutism) reveal that dysfunction in a widespread cortical network including medial and lateral prefrontal and parietal associative areas is associated with a global loss of awareness.[26] Impaired consciousness in epileptic seizures of the temporal lobe was likewise accompanied by a decrease in cerebral blood flow in frontal and parietal association cortex and an increase in midline structures such as the mediodorsal thalamus.[27] Relatively local bilateral injuries to midline (paramedian) subcortical structures can also cause a complete loss of awareness.[28] These structures therefore enable and control brain arousal (as determined by metabolic or electrical activity) and are necessary neural correlates. One such example is the heterogeneous collection of more than two dozen nuclei on each side of the upper brainstem (pons, midbrain and in the posterior hypothalamus), collectively referred to as the reticular activating system (RAS). Their axons project widely throughout the brain. These nuclei – three-dimensional collections of neurons with their own cyto-architecture and neurochemical identity – release distinct neuromodulators such as acetylcholine, noradrenaline/norepinephrine, serotonin, histamine and orexin/hypocretin to control the excitability of the thalamus and forebrain, mediating alternation between wakefulness and sleep as well as general level of behavioral and brain arousal. After such trauma, however, eventually the excitability of the thalamus and forebrain can recover and consciousness can return.[29] Another enabling factor for consciousness are the five or more intralaminar nuclei (ILN) of the thalamus. These receive input from many brainstem nuclei and project strongly, directly to the basal ganglia and, in a more distributed manner, into layer I of much of the neocortex. Comparatively small (1 cm3 or less) bilateral lesions in the thalamic ILN completely knock out all awareness.[30] |
意識の全体的な障害 意識に必要な最小限の神経相関の基準が認められていないため、規則正しい睡眠波移行を示し、動いたり微笑んだりできる可能性がある持続性植物状態の患者 と、意味のある方法で(例えば、差動眼球運動によって)コミュニケーションを取ることができ、意識の兆候を示す最小限の意識状態の患者との区別は、しばし ば困難である。全身麻酔では患者に心理的外傷が生じることはないが、覚醒レベルは臨床的な緊急事態に対応できるものでなければならない。 脳幹と視床にある正中線構造は、脳の覚醒レベルを調節するために必要である。これらの核の多くに小規模な両側病変が生じると、意識が全体的に喪失する。 [24] 重度外傷性脳損傷後の植物状態の患者に、テニスをしているところや自宅の部屋を訪れているところを想像するように指示したところ、血液酸素レベル依存性機 能的磁気共鳴画像法(fMRI)により、脳活動の正常なパターンが示された。[25] このような広範囲の意識障害(運動不能性無言症を含む)を持つ患者の脳画像を比較したところ、 内側および外側前頭前野および頭頂連合野を含む広範囲の皮質ネットワークの機能障害が、意識の全体的な喪失と関連していることが明らかになっている。 [26] 側頭葉のてんかん発作における意識障害も同様に、前頭連合野および頭頂連合野の脳血流の減少と、視床正中背外側部などの正中線構造の増加を伴っている。 [27] また、正中線上(パラメディアン)の皮質下構造に対する比較的限局した両側性の損傷によっても、意識の完全な喪失が引き起こされることがある。[28] これらの構造は、したがって、脳の覚醒を可能にし、制御する(代謝または電気的活動によって決定される)ものであり、必要な神経相関である。その一例とし て、上脳幹(橋、中脳、後部視床下部)のそれぞれに24個以上の核からなる異種混合集団があり、これらは網様体賦活系(RAS)と総称される。それらの軸 索は脳全体に広範囲に広がっている。これらの神経核は、独自の細胞構築と神経化学的同一性を持つニューロンの3次元集合体であり、アセチルコリン、ノルア ドレナリン/ノルエピネフリン、セロトニン、ヒスタミン、オレキシン/ヒポクレチンなどの異なる神経調節物質を放出して、視床および前脳の興奮性を制御 し、覚醒と睡眠の交代、および行動と脳の覚醒の一般的なレベルを調節している。しかし、そのような外傷の後、最終的には視床および前脳の興奮性は回復し、 意識が戻ることもある。[29] 意識を可能にするもう一つの要因は、視床の5つ以上の層内核(ILN)である。これらは多くの脳幹核から入力を受け、直接的に強く大脳基底核に投射し、よ り分散的に大脳新皮質の大部分の層Iに投射する。視床ILNの比較的小さな(1cm3以下)両側病変は、完全にすべての意識を奪う。[30] |
| Forward versus feedback
projections Many actions in response to sensory inputs are rapid, transient, stereotyped, and unconscious.[31] They could be thought of as cortical reflexes and are characterized by rapid and somewhat stereotyped responses that can take the form of rather complex automated behavior as seen, e.g., in complex partial epileptic seizures. These automated responses, sometimes called zombie behaviors,[32] could be contrasted by a slower, all-purpose conscious mode that deals more slowly with broader, less stereotyped aspects of the sensory inputs (or a reflection of these, as in imagery) and takes time to decide on appropriate thoughts and responses. Without such a consciousness mode, a vast number of different zombie modes would be required to react to unusual events. A feature that distinguishes humans from most animals is that we are not born with an extensive repertoire of behavioral programs that would enable us to survive on our own ("physiological prematurity"). To compensate for this, we have an unmatched ability to learn, i.e., to consciously acquire such programs by imitation or exploration. Once consciously acquired and sufficiently exercised, these programs can become automated to the extent that their execution happens beyond the realms of our awareness. Take, as an example, the incredible fine motor skills exerted in playing a Beethoven piano sonata or the sensorimotor coordination required to ride a motorcycle along a curvy mountain road. Such complex behaviors are possible only because a sufficient number of the subprograms involved can be executed with minimal or even suspended conscious control. In fact, the conscious system may actually interfere somewhat with these automated programs.[33] From an evolutionary standpoint it clearly makes sense to have both automated behavioral programs that can be executed rapidly in a stereotyped and automated manner, and a slightly slower system that allows time for thinking and planning more complex behavior. This latter aspect may be one of the principal functions of consciousness. Other philosophers, however, have suggested that consciousness would not be necessary for any functional advantage in evolutionary processes.[34][35] No one has given a causal explanation, they argue, of why it would not be possible for a functionally equivalent non-conscious organism (i.e., a philosophical zombie) to achieve the very same survival advantages as a conscious organism. If evolutionary processes are blind to the difference between function F being performed by conscious organism O and non-conscious organism O*, it is unclear what adaptive advantage consciousness could provide.[36] As a result, an exaptive explanation of consciousness has gained favor with some theorists that posit consciousness did not evolve as an adaptation but was an exaptation arising as a consequence of other developments such as increases in brain size or cortical rearrangement.[37] Consciousness in this sense has been compared to the blind spot in the retina where it is not an adaption of the retina, but instead just a by-product of the way the retinal axons were wired.[38] Several scholars including Pinker, Chomsky, Edelman, and Luria have indicated the importance of the emergence of human language as an important regulative mechanism of learning and memory in the context of the development of higher-order consciousness. It seems possible that visual zombie modes in the cortex mainly use the dorsal stream in the parietal region.[31] However, parietal activity can affect consciousness by producing attentional effects on the ventral stream, at least under some circumstances. The conscious mode for vision depends largely on the early visual areas (beyond V1) and especially on the ventral stream. Seemingly complex visual processing (such as detecting animals in natural, cluttered scenes) can be accomplished by the human cortex within 130–150 ms,[39][40] far too brief for eye movements and conscious perception to occur. Furthermore, reflexes such as the oculovestibular reflex take place at even more rapid time-scales. It is quite plausible that such behaviors are mediated by a purely feed-forward moving wave of spiking activity that passes from the retina through V1, into V4, IT and prefrontal cortex, until it affects motorneurons in the spinal cord that control the finger press (as in a typical laboratory experiment). The hypothesis that the basic processing of information is feedforward is supported most directly by the short times (approx. 100 ms) required for a selective response to appear in IT cells. Conversely, conscious perception is believed to require more sustained, reverberatory neural activity, most likely via global feedback from frontal regions of neocortex back to sensory cortical areas[20] that builds up over time until it exceeds a critical threshold. At this point, the sustained neural activity rapidly propagates to parietal, prefrontal and anterior cingulate cortical regions, thalamus, claustrum and related structures that support short-term memory, multi-modality integration, planning, speech, and other processes intimately related to consciousness. Competition prevents more than one or a very small number of percepts to be simultaneously and actively represented. This is the core hypothesis of the global workspace theory of consciousness.[41][42] In brief, while rapid but transient neural activity in the thalamo-cortical system can mediate complex behavior without conscious sensation, it is surmised that consciousness requires sustained but well-organized neural activity dependent on long-range cortico-cortical feedback. |
予測とフィードバック 感覚入力に対する多くの行動は、迅速かつ一過性で、型にはまった無意識的なものである。[31] これらは大脳皮質の反射と考えられ、迅速かつある程度型にはまった反応によって特徴づけられる。この反応は、例えば複雑部分てんかん発作に見られるよう な、かなり複雑な自動化された行動の形態を取る可能性がある。これらの自動的な反応は、ゾンビ行動と呼ばれることもあるが[32]、より遅く、より汎用的 な意識モードと対照的である。この意識モードは、感覚入力のより広範で定型性の低い側面(またはイメージのようにそれらの反映)をよりゆっくりと処理し、 適切な思考や反応を決定するのに時間を要する。このような意識モードがなければ、異常な事象に対応するには膨大な数の異なるゾンビモードが必要となる。 人間を他の動物と区別する特徴は、人間は自力で生き延びるための行動プログラムの幅広いレパートリーを持って生まれてこないことである(「生理学的未熟 性」)。この欠点を補うために、人間は模倣や探索を通じて、こうしたプログラムを意識的に習得する、つまり学ぶ能力に秀でている。いったん意識的に習得 し、十分に実践すれば、これらのプログラムは自動化され、その実行が意識の及ばない領域で行われるようになる。例えば、ベートーベンのピアノソナタを演奏 する際に発揮される驚くべき細かい運動能力や、曲がりくねった山道をバイクで走るために必要な感覚運動の調整能力などが挙げられる。このような複雑な行動 が可能になるのは、関与するサブプログラムの数が十分にあって、最小限の、あるいは意識的な制御を停止した状態でも実行できるからである。実際には、意識 的なシステムがこれらの自動化されたプログラムに多少干渉している可能性もある。 進化論的な観点から見ると、定型化され自動的に実行される行動プログラムと、より複雑な行動を考える時間や計画を立てる時間を可能にする、やや遅いシステ ムの両方を持つことは明らかに理にかなっている。この後者の側面が意識の主な機能のひとつである可能性がある。しかし、他の哲学者たちは、進化の過程にお いて、意識が機能上の利点をもたらすことはないだろうと主張している。[34][35] 彼らは、機能的に等価な非意識的な生物(すなわち、哲学上のゾンビ)が、意識的な生物と同じ生存上の利点を得ることが不可能である理由について、因果関係 を説明したものは誰もいないと主張している。もし進化の過程が、意識を持つ生物Oと意識を持たない生物O*が機能Fを発揮することの違いを認識しないので あれば、意識がどのような適応上の利点をもたらすのかは不明である。[36] その結果、意識の進化説は、意識は適応として進化するのではなく、脳のサイズの増加や大脳皮質の再編成といった他の進化の結果として生じるエキザプション であると主張する一部の理論家たちに支持されるようになった。[37] この意味での意識は この意味での意識は、網膜の盲点に例えられ、それは網膜の適応ではなく、網膜軸索の配線方法の副産物に過ぎない。[38] ピンカー、チョムスキー、エーデルマン、ルリアを含む複数の学者は、高次意識の発達という文脈において、学習と記憶の重要な調節メカニズムとして、人間の 言語の出現の重要性を指摘している。 大脳皮質における視覚ゾンビモードは主に頭頂葉の背側経路を使用している可能性がある。[31] しかし、少なくともある状況下では、頭頂葉の活動が腹側経路に注意効果を生じさせることで意識に影響を与える可能性もある。視覚に関する意識モードは、初 期視覚野(V1より後)に大きく依存しており、特に腹側経路に依存している。 一見複雑な視覚処理(自然の雑然とした場面で動物を見つけるなど)は、人間の皮質で130~150ミリ秒以内に完了するが、これは眼球運動や意識的な知覚 が起こるにはあまりにも短い時間である。さらに、動眼神経性眼振などの反射は、さらに高速の時間スケールで起こる。このような行動は、網膜からV1、 V4、IT、前頭前野を通り、脊髄の運動ニューロンに影響を与え、指の押し下げを制御する(典型的な実験室での実験のように)純粋なフィードフォワードの スパイク活動の移動波によって媒介される可能性が高い。情報の基本処理がフィードフォワードであるという仮説は、選択的反応が IT 細胞に現れるのに必要な時間が短い(約 100 ミリ秒)という事実によって最も直接的に裏付けられている。 逆に、意識的な知覚には、おそらく新皮質の前頭領域から感覚皮質領域への全体的なフィードバックを通じて、より持続的で反響的な神経活動が必要であると考 えられている。この時点で、持続的な神経活動は急速に頭頂葉、前頭前野、前帯状皮質、視床、クラウストラム、および短期記憶、多感覚統合、計画、言語、お よび意識と密接に関連するその他のプロセスを支える関連構造に伝播する。 競争により、複数の知覚または非常に少数の知覚が同時にかつ能動的に表現されることが妨げられる。 これが意識のグローバルワークスペース理論の中核となる仮説である。[41][42] 簡単に言えば、視床皮質系における急速ではあるが一時的な神経活動は、意識的な感覚なしに複雑な行動を媒介することができるが、意識には、長距離皮質皮質 フィードバックに依存した持続的かつよく組織化された神経活動が必要であると推測されている。 |
| History The neurobiologist Christfried Jakob (1866–1956) argued that the only conditions which must have neural correlates are direct sensations and reactions; these are called "intonations".[citation needed] Neurophysiological studies in animals provided some insights on the neural correlates of conscious behavior. Vernon Mountcastle, in the early 1960s, set up to study this set of problems, which he termed "the Mind/Brain problem", by studying the neural basis of perception in the somatic sensory system. His labs at Johns Hopkins were among the first, along with Edward V.Evarts at NIH, to record neural activity from behaving monkeys. Struck with the elegance of SS Stevens approach of magnitude estimation, Mountcastle's group discovered three different modalities of somatic sensation shared one cognitive attribute: in all cases the firing rate of peripheral neurons was linearly related to the strength of the percept elicited. More recently, Ken H. Britten, William T. Newsome, and C. Daniel Salzman have shown that in area MT of monkeys, neurons respond with variability that suggests they are the basis of decision making about direction of motion. They first showed that neuronal rates are predictive of decisions using signal detection theory, and then that stimulation of these neurons could predictably bias the decision. Such studies were followed by Ranulfo Romo in the somatic sensory system, to confirm, using a different percept and brain area, that a small number of neurons in one brain area underlie perceptual decisions. Other lab groups have followed Mountcastle's seminal work relating cognitive variables to neuronal activity with more complex cognitive tasks. Although monkeys cannot talk about their perceptions, behavioral tasks have been created in which animals made nonverbal reports, for example by producing hand movements. Many of these studies employ perceptual illusions as a way to dissociate sensations (i.e., the sensory information that the brain receives) from perceptions (i.e., how the consciousness interprets them). Neuronal patterns that represent perceptions rather than merely sensory input are interpreted as reflecting the neuronal correlate of consciousness. Using such design, Nikos Logothetis and colleagues discovered perception-reflecting neurons in the temporal lobe. They created an experimental situation in which conflicting images were presented to different eyes (i.e., binocular rivalry). Under such conditions, human subjects report bistable percepts: they perceive alternatively one or the other image. Logothetis and colleagues trained the monkeys to report with their arm movements which image they perceived. Temporal lobe neurons in Logothetis experiments often reflected what the monkeys' perceived. Neurons with such properties were less frequently observed in the primary visual cortex that corresponds to relatively early stages of visual processing. Another set of experiments using binocular rivalry in humans showed that certain layers of the cortex can be excluded as candidates of the neural correlate of consciousness. Logothetis and colleagues switched the images between eyes during the percept of one of the images. Surprisingly the percept stayed stable. This means that the conscious percept stayed stable and at the same time the primary input to layer 4, which is the input layer, in the visual cortex changed. Therefore, layer 4 can not be a part of the neural correlate of consciousness. Mikhail Lebedev and their colleagues observed a similar phenomenon in monkey prefrontal cortex. In their experiments monkeys reported the perceived direction of visual stimulus movement (which could be an illusion) by making eye movements. Some prefrontal cortex neurons represented actual and some represented perceived displacements of the stimulus. Observation of perception related neurons in prefrontal cortex is consistent with the theory of Christof Koch and Francis Crick who postulated that neural correlate of consciousness resides in prefrontal cortex. Proponents of distributed neuronal processing may likely dispute the view that consciousness has a precise localization in the brain. Francis Crick wrote a popular book, "The Astonishing Hypothesis", whose thesis is that the neural correlate for consciousness lies in our nerve cells and their associated molecules. Crick and his collaborator Christof Koch[43] have sought to avoid philosophical debates that are associated with the study of consciousness, by emphasizing the search for "correlation" and not "causation".[needs update] There is much room for disagreement about the nature of this correlate (e.g., does it require synchronous spikes of neurons in different regions of the brain? Is the co-activation of frontal or parietal areas necessary?). The philosopher David Chalmers maintains that a neural correlate of consciousness, unlike other correlates such as for memory, will fail to offer a satisfactory explanation of the phenomenon; he calls this the hard problem of consciousness.[44][45] |
歴史 神経生物学者のクリストフリート・ヤコブ(1866年-1956年)は、神経相関を持つ必要がある唯一の条件は直接的な感覚と反応であると主張した。これ らは「イントネーション」と呼ばれる。[要出典] 動物における神経生理学的研究により、意識的な行動の神経相関に関するいくつかの洞察が得られた。 ヴァーノン・マウントキャッスルは1960年代初頭、体性感覚系における知覚の神経学的基礎を研究することで、この一連の問題の研究に着手し、それを「心 と脳の問題」と名付けた。 マウントキャッスルのジョンズ・ホプキンス大学医学部の研究室は、NIHのエドワード・V・エヴァーツとともに、行動中のサルの神経活動を記録した最初の 研究機関のひとつであった。マウントキャッスル氏のグループは、スティーブンス氏の大きさ推定のアプローチの優雅さに感銘を受け、体性感覚の3つの異なる 様式が1つの認知属性を共有していることを発見した。すなわち、いずれの場合も末梢神経細胞の発火率は、引き起こされた知覚の強さに線形に相関していた。 さらに最近では、ケン・H・ブリテン氏、ウィリアム・T・ニューサム氏、C・ダニエル・サルツマン氏が、サルのMT領域において、神経細胞が変動しながら 反応し、それが運動の方向に関する意思決定の基礎となっていることを示した。彼らはまず、信号検出理論を用いて神経細胞の割合が意思決定を予測できること を示し、次に、これらの神経細胞を刺激することで意思決定を予測可能な形で偏らせることができることを示した。このような研究は、体性感覚系において Ranulfo Romoによって引き継がれ、異なる知覚と脳領域を用いて、ある脳領域の少数の神経細胞が知覚的決定の基礎となっていることを確認した。 他の研究室グループは、Mountcastleの画期的な研究に続き、より複雑な認知課題を用いて、認知変数と神経細胞の活動を関連付けた。サルは知覚に ついて語ることはできないが、動物が非言語的な報告を行う行動課題が考案されている。例えば、手を動かすなどである。これらの研究の多くは、感覚(すなわ ち、脳が受け取る感覚情報)と知覚(すなわち、意識がそれらをどう解釈するか)を切り離す方法として、知覚の錯覚を利用している。単なる感覚入力ではなく 知覚を表すニューロンのパターンは、意識のニューロン相関を反映していると解釈される。 ニコス・ロゴテティス氏とその同僚は、このような設計を用いて、側頭葉における知覚を反映するニューロンを発見した。彼らは、異なる目に相反するイメージ が提示される(すなわち、両眼競合)という実験状況を作り出した。そのような条件下では、被験者は二分覚知覚を報告する。すなわち、一方または他方のイ メージを交互に知覚する。ロゴテティス氏とその同僚は、サルに、知覚したイメージを腕の動きで報告するように訓練した。Logothetisの実験におけ る側頭葉のニューロンは、多くの場合、サルが知覚したものを反映していた。このような特性を持つニューロンは、視覚処理の比較的初期段階に相当する一次視 覚野ではあまり観察されなかった。人間における両眼競合を用いた別の実験では、大脳皮質の特定の層が意識の神経相関の候補から除外できることが示された。 Logothetis氏らは、一方の画像をサルが知覚している間に、両眼の画像を入れ替えた。驚くべきことに知覚は安定したままであった。これは、意識的 な知覚が安定したままであり、同時に視覚皮質における入力層である第4層への一次入力が変化したことを意味する。したがって、第4層は意識の神経相関の一 部であるとは言えない。ミハイル・レベデフ氏と彼の同僚は、サルの前頭前皮質で同様の現象を観察した。彼らの実験では、サルは視覚刺激の移動方向(錯覚で ある可能性もある)を眼球運動によって知覚した。前頭前皮質のいくつかのニューロンは実際の刺激の移動を表し、また、いくつかのニューロンは知覚された刺 激の移動を表した。前頭前皮質における知覚関連ニューロンの観察結果は、意識の神経相関が前頭前皮質に存在するというクリストフ・コッホとフランシス・ク リックの理論と一致している。分散ニューロン処理の支持者たちは、意識が脳の特定の部位に正確に局在するという見解に異論を唱える可能性が高い。 フランシス・クリックは、意識の神経相関は神経細胞とその関連分子にあるという論文で、一般向けに書かれた『驚くべき仮説』を著した。クリックと共同研究 者であるクリストフ・コッホは、意識の研究に関連する哲学的な議論を避けるため、「因果関係」ではなく「相関関係」の追求を強調している。 この相関関係の性質については、多くの意見の相違がある(例えば、脳の異なる領域におけるニューロンの同時スパイクが必要なのか? 前頭葉または頭頂葉の同時活性化が必要なのか?)。哲学者のデビッド・チャルマーズは、意識の神経相関は、記憶などの他の相関とは異なり、現象の満足のい く説明を提供できないと主張している。彼はこれを意識の難問と呼んでいる。[44][45] |
| Animal consciousness Artificial consciousness Binding problem Bridge locus Cognitive map Conceptual space Global workspace theory Hard problem of consciousness Higher-order theories of consciousness Image schema Information-theoretic death Integrated information theory LIDA (cognitive architecture) Models of neural computation Multiple drafts model Münchhausen trilemma Neural coding Neural decoding Neural substrate Philosophy of mind Quantum cognition Quantum mind |
動物意識 人工意識 束縛問題 ブリッジ・ローカス 認知地図 概念空間 グローバルワークスペース理論 意識のハード問題 意識の高次理論 イメージスキーマ 情報理論的死 統合情報理論 LIDA(認知アーキテクチャ 神経計算モデル 複数の草案モデル ミュンヒハウゼンの三重苦 神経コーディング 神経デコーディング 神経基質 心の哲学 量子認知 量子心 |
| https://en.wikipedia.org/wiki/Neural_correlates_of_consciousness |
|
文献
Copyleft, CC, Mitzub'ixi Quq Chi'j, 1996-2099
Copyleft, CC, Mitzub'ixi Quq Chi'j, 1996-2099
![]()
☆ ☆
☆