古人類と現生人類の交配
Interbreeding between archaic and modern humans
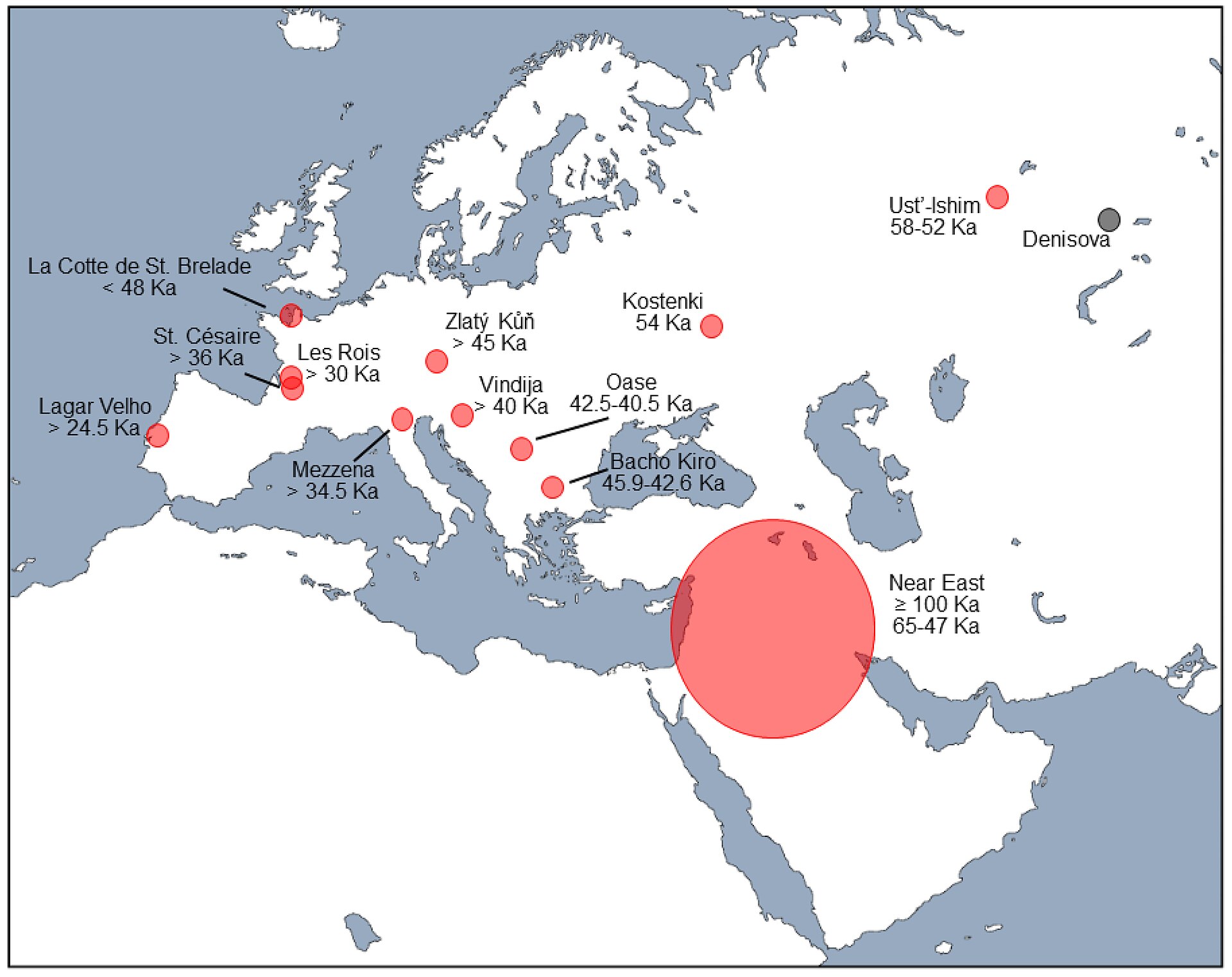
Map
of western Eurasia showing areas and estimated dates of possible
Neanderthal–modern human hybridization (in red) based on fossil samples
from indicated sites
☆ 旧石器人と現生人類の交配(Interbreeding between archaic and modern humans) は、中期旧石器時代と初期上部旧石器時代に起こった。この交雑は、ネアンデルタール人とデニソワ人、そして未同定のホミニンを含むいくつかの独立した出来 事で起こった[2]。 ヨーロッパ、アジア、北アフリカでは、ネアンデルタール人とデニソワ人と現生人類の交雑が数回起こった。現生人類への導入現象は、ネアンデルタール人につ いては約4万7,000~6万5,000年前に、デニソワ人については約4万4,000~5万4,000年前に起こったと推定されている。 ネアンデルタール人由来のDNAは、地域によって顕著に異なるが、現代のほとんどの、あるいはおそらくすべての集団のゲノムから見つかっている。推定値は 異なるが、サハラ以南のアフリカ以外の人々では現代のゲノムの1-4%を占め、サハラ以南のアフリカの人々では全くないか最大0.3%である[3]。西 ユーラシア人に祖先の大部分を持つアフリカの角(エチオピア人など)のクシ族とセム語を話す集団は、ネアンデルタール人由来のDNAを1%程度持っている [4]。 ネアンデルタール人由来のDNAは、東アジア人で最も高く、ヨーロッパ人で中程度、東南アジア人で低い[5]。一部の研究によると、メラネシア人とポリネ シア人は東アジア人とヨーロッパ人の両方と比べても低い[5]。しかし、他の研究では、メラネシア人とネイティブアメリカンでは、ヨーロッパ人よりもネア ンデルタール人の混血が高いことが判明している(東アジア人よりは高くないが)[6]。
| Interbreeding
between archaic and modern humans occurred during the Middle
Paleolithic and early Upper Paleolithic. The interbreeding happened in
several independent events that included Neanderthals and Denisovans,
as well as several unidentified hominins.[2] In Europe, Asia and North Africa, interbreeding between Neanderthals and Denisovans with modern humans took place several times. The introgression events into modern humans are estimated to have happened about 47,000–65,000 years ago with Neanderthals and about 44,000–54,000 years ago with Denisovans. Neanderthal-derived DNA has been found in the genomes of most or possibly all contemporary populations, varying noticeably by region. It accounts for 1–4% of modern genomes for people outside Sub-Saharan Africa, although estimates vary, and either none or up to 0.3% for those in Sub-Saharan Africa.[3] Cushitic and Semitic speaking populations from the Horn of Africa (such as Ethiopians), who derive a large portion of their ancestry from West Eurasians, have ~1% Neanderthal-derived DNA.[4] Neanderthal-derived DNA is highest in East Asians, intermediate in Europeans, and lower in Southeast Asians.[5] According to some research, it is also lower in Melanesians and Polynesians compared to both East Asians and Europeans.[5] However, other research finds higher Neanderthal admixture in Melanesians, as well as in Native Americans, than in Europeans (though not higher than in East Asians).[6] Denisovan-derived ancestry is largely absent from modern populations in Africa, Western Asia and Europe. The highest rates, by far, of Denisovan admixture have been found in Oceanian and some Southeast Asian populations. An estimated 4–6% of the genome of modern Melanesians is derived from Denisovans, but the highest amounts detected thus far are found in the Negrito populations of the Philippines. While some Southeast Asian Negrito populations carry Denisovan admixture, others, such as the Andamanese, have none. In addition, low traces of Denisovan-derived ancestry have been found in mainland Asia, with an elevated Denisovan ancestry in South Asian populations compared to other mainland populations.[7] In Africa, archaic alleles consistent with several independent admixture events in the subcontinent have been found. It is currently unknown who these archaic African hominins were.[5] A 2020 paper found that "despite their very low levels or absence of archaic ancestry, African populations share many Neanderthal and Denisovan variants that are absent from Eurasia, reflecting how a larger proportion of the ancestral human variation has been maintained in Africa."[8] A 2016 paper in the journal Evolutionary Biology argued that introgression of DNA from other lineages enabled humanity to migrate to, and succeed in, numerous new environments, with the resulting hybridization being an essential force in the emergence of modern humans.[9] In December 2023, scientists reported that genes inherited by modern humans from Neanderthals and Denisovans may biologically influence the daily routine of modern humans.[10] |
旧石器人と現生人類の交配は、中期旧石器時代と初期上部旧石器時代に起
こった。この交雑は、ネアンデルタール人とデニソワ人、そして未同定のホミニンを含むいくつかの独立した出来事で起こった[2]。 ヨーロッパ、アジア、北アフリカでは、ネアンデルタール人とデニソワ人と現生人類の交雑が数回起こった。現生人類への導入現象は、ネアンデルタール人につ いては約4万7,000~6万5,000年前に、デニソワ人については約4万4,000~5万4,000年前に起こったと推定されている。 ネアンデルタール人由来のDNAは、地域によって顕著に異なるが、現代のほとんどの、あるいはおそらくすべての集団のゲノムから見つかっている。推定値は 異なるが、サハラ以南のアフリカ以外の人々では現代のゲノムの1-4%を占め、サハラ以南のアフリカの人々では全くないか最大0.3%である[3]。西 ユーラシア人に祖先の大部分を持つアフリカの角(エチオピア人など)のクシ族とセム語を話す集団は、ネアンデルタール人由来のDNAを1%程度持っている [4]。 ネアンデルタール人由来のDNAは、東アジア人で最も高く、ヨーロッパ人で中程度、東南アジア人で低い[5]。 一部の研究によると、メラネシア人とポリネシア人は東アジア人とヨーロッパ人の両方と比べても低い[5]。しかし、他の研究では、メラネシア人とネイティ ブアメリカンでは、ヨーロッパ人よりもネアンデルタール人の混血が高いことが判明している(東アジア人よりは高くないが)[6]。 デニソワ人由来の祖先は、アフリカ、西アジア、ヨーロッパの現代の集団にはほとんど見られない。デニソワ人との混血率が最も高いのは、オセアニアと一部の 東南アジアの集団である。現代のメラネシア人のゲノムの推定4~6%はデニソワ人に由来するが、これまでに検出された中で最も多いのはフィリピンのネグリ ト集団である。東南アジアのネグリト集団にはデニソワ人との混血が見られるものもあるが、アンダマン人のように混血が見られないものもある。さらに、アジ ア本土ではデニソワ人由来の祖先の痕跡は少なく、他の本土の集団と比較して、南アジアの集団ではデニソワ人の祖先の痕跡が高いことが判明している[7]。 アフリカでは、亜大陸におけるいくつかの独立した混血現象と一致する古代の対立遺伝子が見つかっている。2020年に発表された論文によると、「アフリカ の集団は、古代の祖先のレベルが非常に低いか、あるいは存在しないにもかかわらず、ユーラシアには存在しない多くのネアンデルタール人とデニソワ人の変種 を共有しており、これは祖先のヒトの変異のより大きな割合がアフリカで維持されてきたことを反映している」[8]。 Evolutionary Biology』誌に2016年に掲載された論文では、他の系統からのDNAの導入が、人類が多くの新しい環境に移住し、そこで成功することを可能にし、 その結果生じたハイブリダイゼーションが現生人類の出現に不可欠な力であったと論じている[9]。 2023年12月、科学者たちは、現代人がネアンデルタール人やデニソワ人から受け継いだ遺伝子が、現代人の日常生活に生物学的に影響を与えている可能性 があると報告した[10]。 |
| Neanderthals Genetics Main article: Neanderthal genetics Proportion of admixture  Svante Pääbo, Nobel Prize laureate and one of the researchers who published the first sequence of the Neanderthal genome. On 7 May 2010, following the genome sequencing of three Vindija Neanderthals, a draft sequence of the Neanderthal genome was published and revealed that Neanderthals shared more alleles with Eurasian populations (e.g. French, Han Chinese, and Papua New Guinean) than with sub-Saharan African populations (e.g. Yoruba and San).[11] According to the authors Green et al. (2010), the observed excess of genetic similarity is best explained by recent gene flow from Neanderthals to modern humans after the migration out of Africa.[11] They estimated the proportion of Neanderthal-derived ancestry to be 1–4% of the Eurasian genome.[11] Durand et al. (2011) estimated 1–6% Neanderthal ancestry in non-Africans.[12] Prüfer et al. (2013) estimated the proportion to be 1.5–2.1% for non-Africans.[13] Lohse and Frantz (2014) infer a higher rate of 3.4–7.3% in Eurasia.[14] In 2017, Prüfer et al. revised their estimate to 1.8–2.6% for non-Africans outside Oceania.[15] According to a later study by Chen et al. (2020), Africans (specifically, the 1000 Genomes African populations) also have Neanderthal admixture,[16] with this Neanderthal admixture in African individuals accounting for 17 megabases,[16] which is 0.3% of their genome.[3] According to the authors, Africans gained their Neanderthal admixture predominantly from a back-migration by peoples (modern humans carrying Neanderthal admixture) that had diverged from ancestral Europeans (postdating the split between East Asians and Europeans).[16] This back-migration is proposed to have happened about 20,000 years ago.[3] However, some scientists, such as geneticist David Reich, have doubts about how extensive the flow of DNA back to Africa would have been, finding the signal of Neanderthal admixture "really weak".[17] Introgressed genome It has been found that 50% of the Neanderthal genome is present among people in India,[18] and 41% has been found in Icelanders.[19] Previously it was found that about 20% of the Neanderthal genome was found in modern Eurasians,[20] but the figure was also estimated at a third.[21] A 2023 study found an introgession from modern humans to Neanderthals around 250,000 years ago, and estimated that roughly 6% of the Altai Neanderthal genome was inherited from modern humans.[22] Subpopulation admixture rate A higher Neanderthal admixture was found in East Asians than in Europeans,[20][23][24][25][26] which is estimated to be about 20% more introgression into East Asians.[20][23][26] This could possibly be explained by the occurrence of further admixture events in the early ancestors of East Asians after the separation of Europeans and East Asians,[5][20][23][24][26] dilution of Neanderthal ancestry in Europeans by populations with low Neanderthal ancestry from later migrations,[5][23][26] or reduced efficacy of purifying selection in the ancestors of East Asians, due smaller effective population sizes as they migrated to East Asia.[5][25][26] Studies simulating admixture models indicate that a reduced efficacy of purifying selection against Neanderthal alleles in East Asians could not account for the greater proportion of Neanderthal ancestry of East Asians, thus favoring more-complex models involving additional pulses of admixture between Neanderthals and the ancestors of East Asians.[27][28] Such models show a pulse to ancestral Eurasians, followed by separation and an additional pulse to ancestral East Asians.[5] It is observed that there is a small but significant variation of Neanderthal admixture rates within European populations, but no significant variation within East Asian populations.[20] Prüfer et al. (2017) remarked that East Asians carry more Neanderthal DNA (2.3–2.6%) than Western Eurasians (1.8–2.4%).[15] It was later determined by Chen et al. (2020) that East Asians have 8% more Neanderthal ancestry, revised from the previous reports of 20% more Neanderthal ancestry, compared to Europeans.[16] This stems from the fact that Neanderthal ancestry shared with Africans had been masked, because Africans were thought to have no Neanderthal admixture and were therefore used as reference samples.[16] Thus, any overlap in Neanderthal admixture with Africans resulted in an underestimation of Neanderthal admixture in non-Africans and especially in Europeans.[16] The authors give a single pulse of Neanderthal admixture after the out-of-Africa dispersal as the most parsimonious explanation for the enrichment in East Asians, but they add that variation in Neanderthal ancestry may also be attributed to dilution to account for the now-more-modest differences found.[16] As a proportion of the total amount of Neanderthal sequence for each population, 7.2% of the sequence in Europeans is shared exclusively with Africans, while 2% of the sequence in East Asians is shared exclusively with Africans.[16] Genomic analysis suggests that there is a global division in Neanderthal introgression between sub-Saharan African populations and other modern human groups (including North Africans) rather than between African and non-African populations.[29] North African groups share a similar excess of derived alleles with Neanderthals as do non-African populations, whereas sub-Saharan African groups are the only modern human populations that generally did not experience Neanderthal admixture.[30] The Neanderthal genetic signal among North African populations was found to vary depending on the relative quantity of North African, European, Near Eastern and sub-Saharan ancestry. Using F4 ancestry ratio statistical analysis, the Neanderthal inferred admixture was observed to be: highest among the North African populations with highest North African ancestry such as Tunisian Berbers, where it was at the same level or even higher than that of Eurasian populations (100–138%); high among North African populations carrying greater European or Near Eastern admixture, such as groups in North Morocco and Egypt (~60–70%); and lowest among North African populations with greater Sub-Saharan admixture, such as in South Morocco (20%).[31] Quinto et al. (2012) therefore postulate that the presence of this Neanderthal genetic signal in Africa is not due to recent gene flow from Near Eastern or European populations since it is higher among populations bearing indigenous pre-Neolithic North African ancestry.[32] Low but significant rates of Neanderthal admixture has also been observed for the Maasai of East Africa.[33] After identifying African and non-African ancestry among the Maasai, it can be concluded that recent non-African modern human (post-Neanderthal) gene flow was the source of the contribution since around an estimated 30% of the Maasai genome can be traced to non-African introgression from about 100 generations ago.[24] Distance to lineages  Le Moustier Neanderthal skull reconstitution, Neues Museum Berlin.[34] Presenting a high-quality genome sequence of a female Altai Neanderthal, it has been found that the Neanderthal component in non-African modern humans is more related to the Mezmaiskaya Neanderthal (North Caucasus) than to the Altai Neanderthal (Siberia) or the Vindija Neanderthals (Croatia).[13] By high-coverage sequencing the genome of a 50,000-year-old female Vindija Neanderthal fragment, it was later found that the Vindija and Mezmaiskaya Neanderthals did not seem to differ in the extent of their allele-sharing with modern humans.[15] In this case, it was also found that the Neanderthal component in non-African modern humans is more closely related to the Vindija and Mezmaiskaya Neanderthals than to the Altai Neanderthal.[15] These results suggest that a majority of the admixture into modern humans came from Neanderthal populations that had diverged (about 80–100kya) from the Vindija and Mezmaiskaya Neanderthal lineages before the latter two diverged from each other.[15] Analysis of chromosome 21 of the Altai, El Sidrón (Spain), and Vindija Neanderthals indicates that of these three lineages, only the El Sidrón and Vindija Neanderthals display significant rates of gene flow (0.3–2.6%) into modern humans, suggesting that the El Sidrón and Vindija Neanderthals are more closely related than the Altai Neanderthal to the Neanderthals that interbred with modern humans about 47,000–65,000 years ago.[35] Conversely, significant rates of modern human gene flow into Neanderthals occurred—of the three examined lineages—for only the Altai Neanderthal (0.1–2.1%), suggesting that modern human gene flow into Neanderthals mainly took place after the separation of the Altai Neanderthals from the El Sidrón and Vindija Neanderthals that occurred roughly 110,000 years ago.[35] The findings show that the source of modern human gene flow into Neanderthals originated from a population of early modern humans from about 100,000 years ago, predating the out-of-Africa migration of the modern human ancestors of present-day non-Africans.[35] Mitochondrial DNA and Y chromosome No evidence of Neanderthal mitochondrial DNA has been found in modern humans.[36][37][38] This suggests that successful Neanderthal admixture happened in pairings with Neanderthal males and modern human females.[39][40] Possible hypotheses are that Neanderthal mitochondrial DNA had detrimental mutations that led to the extinction of carriers, that the hybrid offspring of Neanderthal mothers were raised in Neanderthal groups and became extinct with them, or that female Neanderthals and male Sapiens did not produce fertile offspring.[39] However, the hypothesized incompatibility between Neanderthals and modern humans is contested by findings that suggest that the Y chromosome of Neanderthals was replaced by an extinct lineage of the modern human Y chromosome, which introgressed into Neanderthals between 100,000 and 370,000 years ago.[41] Furthermore, the study concludes that the replacement of the Y chromosomes and mitochondrial DNA in Neanderthals after gene flow from modern humans is highly plausible given the increased genetic load in Neanderthals relative to modern humans.[41] As shown in an interbreeding model produced by Neves and Serva (2012), the Neanderthal admixture in modern humans may have been caused by a very low rate of interbreeding between modern humans and Neanderthals, with the exchange of one pair of individuals between the two populations in about every 77 generations.[42] This low rate of interbreeding would account for the absence of Neanderthal mitochondrial DNA from the modern human gene pool as found in earlier studies, as the model estimates a probability of only 7% for a Neanderthal origin of both mitochondrial DNA and Y chromosome in modern humans.[42] Reduced contribution There are large genomic regions with strongly reduced Neanderthal contribution in modern humans due to negative selection,[20][25] partly caused by hybrid male infertility.[25] These regions were most-pronounced on the X chromosome, with fivefold lower Neanderthal ancestry compared to autosomes.[5][25] They also contained relatively high numbers of genes specific to testes.[25] This means that modern humans have relatively few Neanderthal genes that are located on the X chromosome or expressed in the testes, suggesting male infertility as a probable cause.[25] It may be partly affected by hemizygosity of X chromosome genes in males.[5] Deserts of Neanderthal sequences may also be caused by genetic drift involving intense bottlenecks in the modern human population and background selection as a result of strong selection against deleterious Neanderthal alleles.[5] The overlap of many deserts of Neanderthal and Denisovan sequences suggests that repeated loss of archaic DNA occur at specific loci.[5] It has also been shown that Neanderthal ancestry has been selected against in conserved biological pathways, such as RNA processing.[25] Consistent with the hypothesis that purifying selection has reduced Neanderthal contribution in present-day modern human genomes, Upper Paleolithic Eurasian modern humans (such as the Tianyuan modern human) carry more Neanderthal DNA (about 4–5%) than present-day Eurasian modern humans (about 1–2%).[43] Rates of selection against Neanderthal sequences varied for European and Asian populations.[5] |
ネアンデルタール人 遺伝学 主な記事 ネアンデルタール人の遺伝学 混血の割合 ノーベル賞受賞者で、ネアンデルタール人のゲノム配列を初めて発表した研究者の一人、スヴァンテ・ペーボ。 2010年5月7日、3頭のヴィンディヤ・ネアンデルタール人のゲノム配列決定を受けて、ネアンデルタール人ゲノムのドラフト配列が発表され、ネアンデル タール人はサハラ以南のアフリカ人集団(ヨルバ人やサン人など)よりもユーラシア人集団(フランス人、漢民族、パプアニューギニア人など)と対立遺伝子を 多く共有していることが明らかになった。 Green et al. (2010)によれば、観察された過剰な遺伝的類似性は、アフリカから移動したネアンデルタール人から現生人類への最近の遺伝子の流れによって最もよく説 明される[11]。 [11]彼らはネアンデルタール人由来の祖先の割合をユーラシアゲノムの1-4%と推定した[11]。Durandら(2011)は非アフリカ人のネアン デルタール人祖先の割合を1-6%と推定した[12]。 13] Lohse and Frantz (2014)はユーラシア大陸では3.4-7.3%とより高い割合を推測している[14]。2017年にPrüferらはオセアニア以外の非アフリカ人に ついて1.8-2.6%と推定を修正した[15]。 Chenらによる後の研究(2020年)によると、アフリカ人(具体的には1000 Genomesのアフリカ人集団)にもネアンデルタール人との混血があり[16]、アフリカ人個体におけるこのネアンデルタール人との混血は17メガベー ス[16]、ゲノムの0.3%を占めている。 [3] 著者らによれば、アフリカ人がネアンデルタール人との混血を獲得したのは、祖先であるヨーロッパ人から分岐した民族(ネアンデルタール人との混血を持つ現 生人類)が(東アジア人とヨーロッパ人の分裂後に)逆移動してきたことが主因である。 [しかし、遺伝学者のデイヴィッド・ライヒのように、ネアンデルタール人との混血のシグナルは「実に弱い」[17]とし、アフリカへのDNAの逆流がどれ ほど広範なものであったかを疑問視する科学者もいる。 移入されたゲノム ネアンデルタール人ゲノムの50%がインドの人々に存在し[18]、41%がアイスランド人に存在することが判明している[19]。以前はネアンデルター ル人ゲノムの約20%が現代のユーラシア人に存在することが判明していたが[20]、この数字は3分の1とも推定されていた[21]。 亜集団混血率 ネアンデルタール人との混血率はヨーロッパ人よりも東アジア人の方が高く[20][23][24][25][26]、東アジア人への導入率は約20%高い と推定されている。 [20][23][26]これはおそらく,ヨーロッパ人と東アジア人が分離した後,東アジア人の初期の祖先においてさらなる混血事象が発生したこと, [5][20][23][24][26]後の移住によるネアンデルタール人の祖先が少ない集団によってヨーロッパ人のネアンデルタール人の祖先が希釈され たこと,[5][23][26]あるいは,東アジア人の祖先が東アジアに移住するにつれて有効な集団のサイズが小さくなったために,東アジア人の祖先にお ける純化選択の有効性が低下したことによって説明できるかもしれない。 [5][25][26]混血モデルをシミュレートした研究によると、東アジア人におけるネアンデルタール人の対立遺伝子に対する純化選択の有効性が低下し たのでは、東アジア人の祖先がネアンデルタール人である割合が高いことを説明することはできず、ネアンデルタール人と東アジア人の祖先との間の混血の追加 的なパルスを含む、より複雑なモデルが好まれる。 [27][28]そのようなモデルは、祖先ユーラシア人へのパルス、その後の分離、祖先東アジア人への追加パルスを示している[5]。ヨーロッパの集団内 ではネアンデルタール人の混血率にわずかではあるが有意な変動があることが観察されているが、東アジアの集団内では有意な変動はない[20]。 その後、Chenら(2020)によって、東アジア人はヨーロッパ人と比較して、ネアンデルタール人の祖先を20%多く持っているという以前の報告から修 正され、ネアンデルタール人の祖先を8%多く持っていると決定された。 [16]これはアフリカ人と共有されるネアンデルタール人の祖先が、アフリカ人にはネアンデルタール人との混血がないと考えられていたため、参照サンプル として使用され、マスクされていたことに起因する。 [16] 著者らは、東アジア人におけるネアンデルタール人の混血の最も穏当な説明として、アフリカ外への拡散後のネアンデルタール人の混血の単一パルスを挙げてい るが、ネアンデルタール人の祖先のばらつきは希釈に起因している可能性もあり、現在ではより緩やかな違いが見つかっていると付け加えている。 [16]各集団のネアンデルタール人の塩基配列の総量の割合として、ヨーロッパ人の塩基配列の7.2%はアフリカ人とだけ共有されており、東アジア人の塩 基配列の2%はアフリカ人とだけ共有されている[16]。 ゲノム解析によると、ネアンデルタール人の導入は、アフリカ人と非アフリカ人集団の間ではなく、サハラ以南のアフリカ人集団と他の現生人類集団(北アフリ カ人を含む)の間で世界的に分断されている。 [29]北アフリカの集団は非アフリカの集団と同様にネアンデルタール人に由来する対立遺伝子の過剰を共有しているが、サハラ以南のアフリカの集団は一般 的にネアンデルタール人との混血を経験しなかった唯一の現生人類の集団である[30]。北アフリカの集団間のネアンデルタール人の遺伝的シグナルは、北ア フリカ、ヨーロッパ、近東、サハラ以南の祖先の相対的な量によって異なることが判明した。F4祖先比率統計分析を用いると、ネアンデルタール人による混血 は以下のように観察された: チュニジアのベルベル人のような北アフリカの祖先が最も多い集団では最も高く、ユーラシアの集団と同じかそれ以上であった(100-138%)。北モロッ コやエジプトの集団のように、ヨーロッパや近東の混血が多い北アフリカの集団では高く(60-70%)、南モロッコのようなサハラ以南の混血が多い北アフ リカの集団では最も低かった(20%)。 [31] Quinto et al. (2012)は、ネアンデルタール人の遺伝的シグナルがアフリカに存在するのは、近東やヨーロッパの集団からの最近の遺伝子流入によるものではないとして いる。 [33]。マサイ族におけるアフリカ系と非アフリカ系の祖先を同定した結果、最近の非アフリカ系現生人類(ポストネアンデルタール人)の遺伝子フローが寄 与していると結論づけられる。 系統までの距離 ル・ムスティエ・ネアンデルタール人の頭蓋骨の復元、ベルリン・ノイミュージアム[34]。 アルタイ・ネアンデルタール人(女性)の高品質なゲノム配列から、非アフリカ現生人類におけるネアンデルタール人の構成要素は、アルタイ・ネアンデルター ル人(シベリア)やヴィンディヤ・ネアンデルタール人(クロアチア)よりもメズマイスカヤ・ネアンデルタール人(北コーカサス)に近縁であることが判明し た[13]。 [ヴィンディヤ・ネアンデルタール人の5万年前の女性のゲノムをハイカバレッジシークエンシングした結果、ヴィンディヤ・ネアンデルタール人とメズマイス カヤ・ネアンデルタール人は、現代人との対立遺伝子の共有の程度に違いがないことが後に判明した。 [この場合、アフリカ以外の現生人類におけるネアンデルタール人の構成要素は、アルタイ・ネアンデルタール人よりもヴィンディヤ・ネアンデルタール人とメ ズマイスカヤ・ネアンデルタール人に近縁であることも判明した[15]。これらの結果から、現生人類への混血の大部分は、ヴィンディヤ・ネアンデルタール 人とメズマイスカヤ・ネアンデルタール人の系統が互いに分岐する前に、ヴィンディヤ・ネアンデルタール人とメズマイスカヤ・ネアンデルタール人の系統から 分岐(約80-100kya)したネアンデルタール人の集団からもたらされたことが示唆される[15]。 アルタイ人、エル・シドロン人(スペイン)、ヴィンディージャ・ネアンデルタール人の21番染色体を分析した結果、これら3つの系統のうち、エル・シドロ ン人とヴィンディージャ・ネアンデルタール人だけが現代人への有意な遺伝子流入率(0.3-2. このことは、エル・シドロンとヴィンディヤ・ネアンデルタール人がアルタイ・ネアンデルタール人よりも、約4万7,000~6万5,000年前に現生人類 と交雑したネアンデルタール人に近縁であることを示唆している[35]。逆に、ネアンデルタール人への現生人類の遺伝子流入は、調査した3つの系統の中で アルタイ・ネアンデルタール人だけが有意な割合(0. 1-2.1%)、ネアンデルタール人への現生人類の遺伝子流入は、主におよそ11万年前に起こったアルタイ・ネアンデルタール人とエル・シドロン・ネアン デルタール人およびヴィンディジャ・ネアンデルタール人との分離の後に起こったことを示唆している[35]。この発見は、ネアンデルタール人への現生人類 の遺伝子流入の源は、現在の非アフリカ人の現生人類の祖先のアフリカ外への移動よりも前の、およそ10万年前の初期現生人類の集団に由来することを示して いる[35]。 ミトコンドリアDNAとY染色体 ネアンデルタール人のミトコンドリアDNAの証拠は現生人類には見つかっていない[36][37][38]。このことは、ネアンデルタール人の男性と現生 人類の女性とのペアリングにおいて、ネアンデルタール人の混血が成功したことを示唆している。 [39][40]考えられる仮説は、ネアンデルタール人のミトコンドリアDNAに有害な変異があり、それが保因者の絶滅につながった、ネアンデルタール人 の母親のハイブリッド子孫がネアンデルタール人の集団で育てられ、彼らと共に絶滅した、あるいはネアンデルタール人のメスとサピエンスのオスは繁殖可能な 子孫を残せなかった、というものである。 [39]しかし、ネアンデルタール人と現生人類の間の非互換性という仮説は、ネアンデルタール人のY染色体が、10万年から37万年前の間にネアンデル タール人に導入された現生人類のY染色体の絶滅系統に置き換えられたことを示唆する調査結果によって否定されている。 [41]さらに、ネアンデルタール人のY染色体とミトコンドリアDNAが、現生人類からの遺伝子流入後に置換されたことは、現生人類と比較してネアンデル タール人の遺伝的負荷が増加していることを考えると、非常にもっともらしいと結論付けている[41]。 Neves and Serva (2012)が作成した交雑モデルで示されているように、現生人類におけるネアンデルタール人との混血は、現生人類とネアンデルタール人との間の交雑率が 非常に低く、約77世代に1組の個体が2つの集団間で交換されたことによって引き起こされた可能性がある[42]。 [42]。この交雑率の低さは、以前の研究で発見されたように、現生人類の遺伝子プールからネアンデルタール人のミトコンドリアDNAが検出されないこと を説明するものである。 寄与の減少 ネアンデルタール人の祖先は、常染色体と比較して5倍低い[5][25]。 [5][25]また、精巣に特異的な遺伝子を比較的多く含んでいた。[25]このことは、現生人類はX染色体上に位置する、あるいは精巣で発現するネアン デルタール人の遺伝子を比較的少なく持っていることを意味し、男性不妊が原因である可能性が高いことを示唆している。[25]男性におけるX染色体遺伝子 の半接合性が一部影響している可能性がある。 ネアンデルタール人の配列の砂漠は、現代人の集団における激しいボトルネックと、ネアンデルタール人の有害な対立遺伝子に対する強い淘汰の結果としての背 景淘汰を含む遺伝的ドリフトによっても引き起こされる可能性がある[5]。ネアンデルタール人とデニソワ人の配列の砂漠の多くが重なっていることから、特 定の遺伝子座で古代のDNAの喪失が繰り返されていることが示唆されている[5]。 また、ネアンデルタール人の祖先は、RNAプロセシングのような保存された生物学的経路において淘汰されていることも示されている[25]。 純化選択が現生人類のゲノムにおけるネアンデルタール人の寄与を減少させたという仮説と一致して、上旧石器時代のユーラシア現生人類(天元現生人類など) は、現生ユーラシア現生人類(約1-2%)よりも多くのネアンデルタール人DNA(約4-5%)を保持している[43]。 ネアンデルタール人の配列に対する選択の割合は、ヨーロッパとアジアの集団で異なっていた[5]。 |
 |
|
| Changes in modern humans Further information: Recent human evolution In Eurasia, modern humans have adaptive sequences introgressed from archaic humans, which provided a source of advantageous genetic variants that are adapted to local environments and a reservoir for additional genetic variation.[5] Adaptive introgression from Neanderthals has targeted genes involved with keratin filaments, sugar metabolism, muscle contraction, body fat distribution, enamel thickness, and oocyte meiosis, as well as brain size and functioning.[44] There are signals of positive selection, as the result of adaptation to diverse habitats, in genes involved with variation in skin pigmentation and hair morphology.[44] In the immune system, introgressed variants have heavily contributed to the diversity of immune genes, of which there's an enrichment of introgressed alleles that suggest a strong positive selection.[44] Genes affecting keratin were found to have been introgressed from Neanderthals into modern humans (shown in East Asians and Europeans), suggesting that these genes gave a morphological adaptation in skin and hair to modern humans to cope with non-African environments.[20][25] This is likewise for several genes involved in medical-relevant phenotypes, such as those affecting systemic lupus erythematosus, primary biliary cirrhosis, Crohn's disease, optic disk size, smoking behavior, interleukin 18 levels, and diabetes mellitus type 2.[25] Researchers found Neanderthal introgression of 18 genes—several of which are related to UV-light adaptation—within the chromosome 3p21.31 region (HYAL region) of East Asians.[45] The introgressive haplotypes were positively selected in only East Asian populations, rising steadily from 45,000 years BP until a sudden increase of growth rate around 5,000 to 3,500 years BP.[45] They occur at very high frequencies among East Asian populations in contrast to other Eurasian populations (e.g. European and South Asian populations).[45] The findings also suggests that this Neanderthal introgression occurred within the ancestral population shared by East Asians and Native Americans.[45] Evans et al. (2006) had previously suggested that a group of alleles collectively known as haplogroup D of microcephalin, a critical regulatory gene for brain volume, originated from an archaic human population.[46] The results show that haplogroup D introgressed 37,000 years ago (based on the coalescence age of derived D alleles) into modern humans from an archaic human population that separated 1.1 million years ago (based on the separation time between D and non-D alleles), consistent with the period when Neanderthals and modern humans co-existed and diverged respectively.[46] The high frequency of the D haplogroup (70%) suggest that it was positively selected for in modern humans.[46] The distribution of the D allele of microcephalin is high outside Africa but low in sub-Saharan Africa, which further suggest that the admixture event happened in archaic Eurasian populations.[46] This distribution difference between Africa and Eurasia suggests that the D allele originated from Neanderthals according to Lari et al. (2010), but they found that a Neanderthal individual from the Mezzena Rockshelter (Monti Lessini, Italy) was homozygous for an ancestral allele of microcephalin, thus providing no support that Neanderthals contributed the D allele to modern humans and also not excluding the possibility of a Neanderthal origin of the D allele.[47] Green et al. (2010), having analyzed the Vindija Neanderthals, also could not confirm a Neanderthal origin of haplogroup D of the microcephalin gene.[11] It has been found that HLA-A*02, A*26/*66, B*07, B*51, C*07:02, and C*16:02 of the immune system were contributed from Neanderthals to modern humans.[48] After migrating out of Africa, modern humans encountered and interbred with archaic humans, which was advantageous for modern humans in rapidly restoring HLA diversity and acquiring new HLA variants that are better adapted to local pathogens.[48] It is found that introgressed Neanderthal genes exhibit cis-regulatory effects in modern humans, contributing to the genomic complexity and phenotype variation of modern humans.[49] Looking at heterozygous individuals (carrying both Neanderthal and modern human versions of a gene), the allele-specific expression of introgressed Neanderthal alleles was found to be significantly lower in the brain and testes relative to other tissues.[5][49] In the brain, this was most pronounced at the cerebellum and basal ganglia.[49] This downregulation suggests that modern humans and Neanderthals possibly experienced a relative higher rate of divergence in these specific tissues.[49] Furthermore, correlating the genotypes of introgressed Neanderthal alleles with the expression of nearby genes, it is found that archaic alleles contribute proportionally more to variation in expression than nonarchaic alleles.[5] Neanderthal alleles affect expression of the immune genes OAS1/2/3 and TLR1/6/10, which can be specific to cell type and is influenced by environmental stimuli.[5] Studying the high-coverage female Vindija Neanderthal genome, Prüfer et al. (2017) identified several Neanderthal-derived gene variants, including those that affect levels of LDL cholesterol and vitamin D, and that influence eating disorders, visceral fat accumulation, rheumatoid arthritis, schizophrenia, as well as responses to antipsychotic drugs.[15] Examining European modern humans in regards to the Altai Neanderthal genome in high-coverage, results show that Neanderthal admixture is associated with several changes in cranium and underlying brain morphology, suggesting changes in neurological function through Neanderthal-derived genetic variation.[50] Neanderthal admixture is associated with an expansion of the posterolateral area of the modern human skull, extending from the occipital and inferior parietal bones to bilateral temporal locales.[50] In regards to modern human brain morphology, Neanderthal admixture is positively correlated with an increase in sulcal depth for the right intraparietal sulcus and an increase in cortical complexity for the early visual cortex of the left hemisphere.[50] Neanderthal admixture is also positively correlated with an increase in white and gray matter volume localized to the right parietal region adjacent to the right intraparietal sulcus.[50] In the area overlapping the primary visual cortex gyrification in the left hemisphere, Neanderthal admixture is positively correlated with gray matter volume.[50] The results also show evidence for a negative correlation between Neanderthal admixture and white matter volume in the orbitofrontal cortex.[50] In Papuans, Neanderthal genetic variants are found in highest frequency in genes expressed in the brain, whereas Denisovan DNA has the highest frequency in genes expressed in bones and other tissues.[51] A Neanderthal allele inherited by modern humans, SNP rs3917862, is with associated with hypercoagulability. This can be harmful, but women lacking the allele are 0.1% more likely to die in childbirth.[52] In December 2023, scientists reported that genes inherited by modern humans from Neanderthals and Denisovans may biologically influence the daily routine of modern humans.[10] Population substructure theory Although less parsimonious than recent gene flow, the observation may have been due to ancient population sub-structure in Africa, causing incomplete genetic homogenization within modern humans when Neanderthals diverged while early ancestors of Eurasians were still more closely related to Neanderthals than those of Africans were to Neanderthals.[11] On the basis of allele frequency spectrum, it was shown that the recent admixture model had the best fit to the results while the ancient population sub-structure model had no fit—demonstrating that the best model was a recent admixture event that was preceded by a bottleneck event among modern humans – thus confirming recent admixture as the most parsimonious and plausible explanation for the observed excess of genetic similarities between modern non-African humans and Neanderthals.[53] On the basis of linkage disequilibrium patterns, a recent admixture event is likewise confirmed by the data.[54] From the extent of linkage disequilibrium, it was estimated that the last Neanderthal gene flow into early ancestors of Europeans occurred 47,000–65,000 years BP.[54] In conjunction with archaeological and fossil evidence, the gene flow is thought likely to have occurred somewhere in Western Eurasia, possibly the Middle East.[54] Through another approach—using one genome each of a Neanderthal, Eurasian, African, and chimpanzee (outgroup), and dividing it into non-recombining short sequence blocks—to estimate genome-wide maximum-likelihood under different models, an ancient population sub-structure in Africa was ruled out and a Neanderthal admixture event was confirmed.[14] |
現代人の変化 さらに詳しい情報 最近の人類の進化 ユーラシア大陸では、現生人類は古人類から導入された適応配列を持っており、現地の環境に適応した有利な遺伝的変異の供給源となり、さらなる遺伝的変異の 貯蔵庫となった。ネアンデルタール人からの適応的導入は、ケラチンフィラメント、糖代謝、筋肉収縮、体脂肪分布、エナメル質の厚さ、卵母細胞の減数分裂に 関わる遺伝子や、脳の大きさと機能を標的としている。 [44] 皮膚の色素沈着や毛の形態の変化に関わる遺伝子には、多様な生息環境に適応した結果、正の選択のシグナルが見られる。免疫系では、導入された変異体が免疫 遺伝子の多様性に大きく寄与しており、そのうち強い正の選択を示唆する導入された対立遺伝子が濃縮されている。 ケラチンに影響する遺伝子がネアンデルタール人から現生人類に導入されたことが判明し(東アジア人とヨーロッパ人に見られる)、これらの遺伝子が現生人類 に非アフリカ的環境に対処するための皮膚と毛髪の形態的適応を与えたことを示唆している。 [このことは、全身性エリテマトーデス、原発性胆汁性肝硬変、クローン病、視神経円板の大きさ、喫煙行動、インターロイキン18レベル、2型糖尿病に影響 を及ぼす遺伝子など、医学的に関連する表現型に関与するいくつかの遺伝子についても同様である[25]。 研究者らは、東アジア人の染色体3p21.31領域(HYAL領域)に、紫外線適応に関連する18の遺伝子がネアンデルタール人に導入されていることを発 見した[45]。この導入ハプロタイプは、東アジアの集団でのみ正選択されており、45,000年BPから5,000年から3,500年BPの頃に急激に 増加するまで、着実に増加していた。 [45]これらのハプロタイプは、他のユーラシアの集団(ヨーロッパや南アジアの集団など)とは対照的に、東アジアの集団では非常に高い頻度で出現してい る[45]。この発見はまた、このネアンデルタール人の導入が、東アジア人とアメリカ先住民が共有する祖先集団内で起こったことを示唆している[45]。 Evansら(2006年)は以前、脳容積の重要な調節遺伝子であるマイクロセファリンのハプログループDと総称される対立遺伝子群が、古代の人類集団か ら発生したことを示唆していた[46]。この結果は、ハプログループDが37,000年前(由来するD対立遺伝子の合体年齢に基づく)に、110万年前に 分離した古代の人類集団から現生人類に導入されたことを示している。 これはネアンデルタール人と現生人類がそれぞれ共存し、分岐した時期と一致する[46]。Dハプログループの頻度が高い(70%)ことから、現生人類にお いてDハプログループが正に選択されたことが示唆される。 [46]マイクロセファリンのD対立遺伝子の分布は、アフリカ以外では高いが、サハラ以南のアフリカでは低いことから、混血はユーラシア大陸の古代の集団 で起こったことが示唆されている[46]。(2010)によれば、D対立遺伝子はネアンデルタール人由来であることを示唆しているが、彼らは Mezzena Rockshelter(イタリア、Monti Lessini)から出土したネアンデルタール人個体がマイクロセファリンの祖先対立遺伝子に対してホモ接合であることを発見している。 [47]ヴィンディヤのネアンデルタール人を分析したGreenら(2010年)も、ミクロセファリン遺伝子のハプログループDのネアンデルタール人起源 を確認できなかった[11]。 免疫系のHLA-A*02、A*26/*66、B*07、B*51、C*07:02、C*16:02はネアンデルタール人から現生人類に寄与されたことが 判明している[48]。アフリカから移動した現生人類は古人類と遭遇し交雑したが、これは現生人類にとってHLAの多様性を急速に回復させ、現地の病原体 により適応した新しいHLA変異体を獲得する上で有利であった[48]。 ヘテロ接合体(ある遺伝子のネアンデルタール人バージョンと現代人バージョンの両方を持つ)を見ると、導入されたネアンデルタール人対立遺伝子の対立遺伝 子特異的発現は、他の組織に比べて脳と精巣で有意に低いことがわかった。 [この発現低下は、現代人とネアンデルタール人がこれらの特定の組織において相対的に高い分岐率を経験した可能性を示唆している[49]。 さらに、導入されたネアンデルタール人の対立遺伝子の遺伝子型と近傍遺伝子の発現を相関させると、古代の対立遺伝子は非古代の対立遺伝子よりも発現の変動 に比例して大きく寄与していることがわかった[5]。ネアンデルタール人の対立遺伝子は、免疫遺伝子OAS1/2/3とTLR1/6/10の発現に影響を 与えており、これは細胞のタイプに特異的であり、環境刺激の影響を受ける可能性がある[5]。 高カバレッジの女性Vindijaネアンデルタール人ゲノムを研究しているPrüferら(2017年)は、LDLコレステロールやビタミンDのレベルに 影響するもの、摂食障害、内臓脂肪蓄積、関節リウマチ、統合失調症、さらには抗精神病薬への反応に影響するものなど、ネアンデルタール人由来の遺伝子変異 をいくつか同定した[15]。 アルタイ・ネアンデルタール人ゲノムをハイカバレッジでヨーロッパ現生人類を調査した結果、ネアンデルタール人との混血が頭蓋とその下にある脳の形態にお けるいくつかの変化と関連していることが示され、ネアンデルタール人由来の遺伝的変異を通じて神経機能に変化が生じていることが示唆された。 [50] 現代人の脳の形態に関して、ネアンデルタール人との混血は、右頭頂内溝の溝の深さの増加、および左半球の初期視覚野の皮質の複雑さの増加と正の相関があ る。 [50] また、左半球の一次視覚野回旋に重なる領域では、ネアンデルタール人との混血は灰白質容積と正の相関がある[50]。この結果は、ネアンデルタール人との 混血と眼窩前頭皮質の白質容積との間に負の相関があることを示す証拠でもある[50]。 パプア人において、ネアンデルタール人の遺伝子変異は脳で発現する遺伝子で最も高い頻度で見つかっているのに対し、デニソワ人のDNAは骨や他の組織で発現する遺伝子で最も高い頻度で見つかっている[51]。 現代人に受け継がれるネアンデルタール人の対立遺伝子であるSNP rs3917862は、凝固性亢進と関連している。これは有害である可能性があるが、この対立遺伝子を持たない女性は出産時に死亡する可能性が0.1%高い[52]。 2023年12月、科学者たちは、現代人がネアンデルタール人やデニソワ人から受け継いだ遺伝子が、現代人の日常生活に生物学的に影響を与えている可能性があると報告した[10]。 集団下部構造理論 最近の遺伝子の流れよりは単純化できないが、この観測はアフリカにおける古代の集団下部構造によるものであり、ユーラシア人の初期の祖先がネアンデルター ル人とアフリカ人の祖先よりもネアンデルタール人にまだ近縁であった間にネアンデルタール人が分岐した際に、現生人類内で不完全な遺伝的均質化を引き起こ した可能性がある。 [11]対立遺伝子頻度スペクトルに基づくと、最近の混血モデルが結果に対して最も適合していることが示されたが、古代の集団亜構造モデルは適合しなかっ た-このことは、最良のモデルが現生人類の間でボトルネック事象に先行された最近の混血事象であることを実証している-したがって、現代の非アフリカ系人 類とネアンデルタール人との間で観察された遺伝的類似性の過剰に対する最も簡潔でもっともらしい説明が最近の混血であることが確認された。 [連鎖不平衡の範囲から、ネアンデルタール人からヨーロッパ人の初期祖先への最後の遺伝子流入は、47,000-65,000年前に起こったと推定されて いる[54]。 [54] 別のアプローチ-ネアンデルタール人、ユーラシア人、アフリカ人、チンパンジー(アウトグループ)のゲノムを1つずつ使用し、それを非組み合わせ短配列ブ ロックに分割して、異なるモデルの下でゲノム全体の最尤度を推定する-によって、アフリカにおける古代の集団亜構造は除外され、ネアンデルタール人の混血 イベントが確認された[14]。 |
| Morphology The early Upper Paleolithic burial remains of a modern human child from Abrigo do Lagar Velho (Portugal) features traits that indicate Neanderthal interbreeding with modern humans dispersing into Iberia.[55] Considering the dating of the burial remains (24,500 years BP) and the persistence of Neanderthal traits long after the transitional period from a Neanderthal to a modern human population in Iberia (28,000–30,000 years BP), the child may have been a descendant of an already heavily admixed population.[55] The remains of an early Upper Paleolithic modern human from Peștera Muierilor (Romania) of 35,000 years BP shows a morphological pattern of European early modern humans, but possesses archaic or Neanderthal features, suggesting European early modern humans interbreeding with Neanderthals.[56] These features include a large interorbital breadth, a relatively flat superciliary arches, a prominent occipital bun, an asymmetrical and shallow mandibular notch shape, a high mandibular coronoid processus, the relative perpendicular mandibular condyle to notch crest position, and a narrow scapular glenoid fossa.[56] 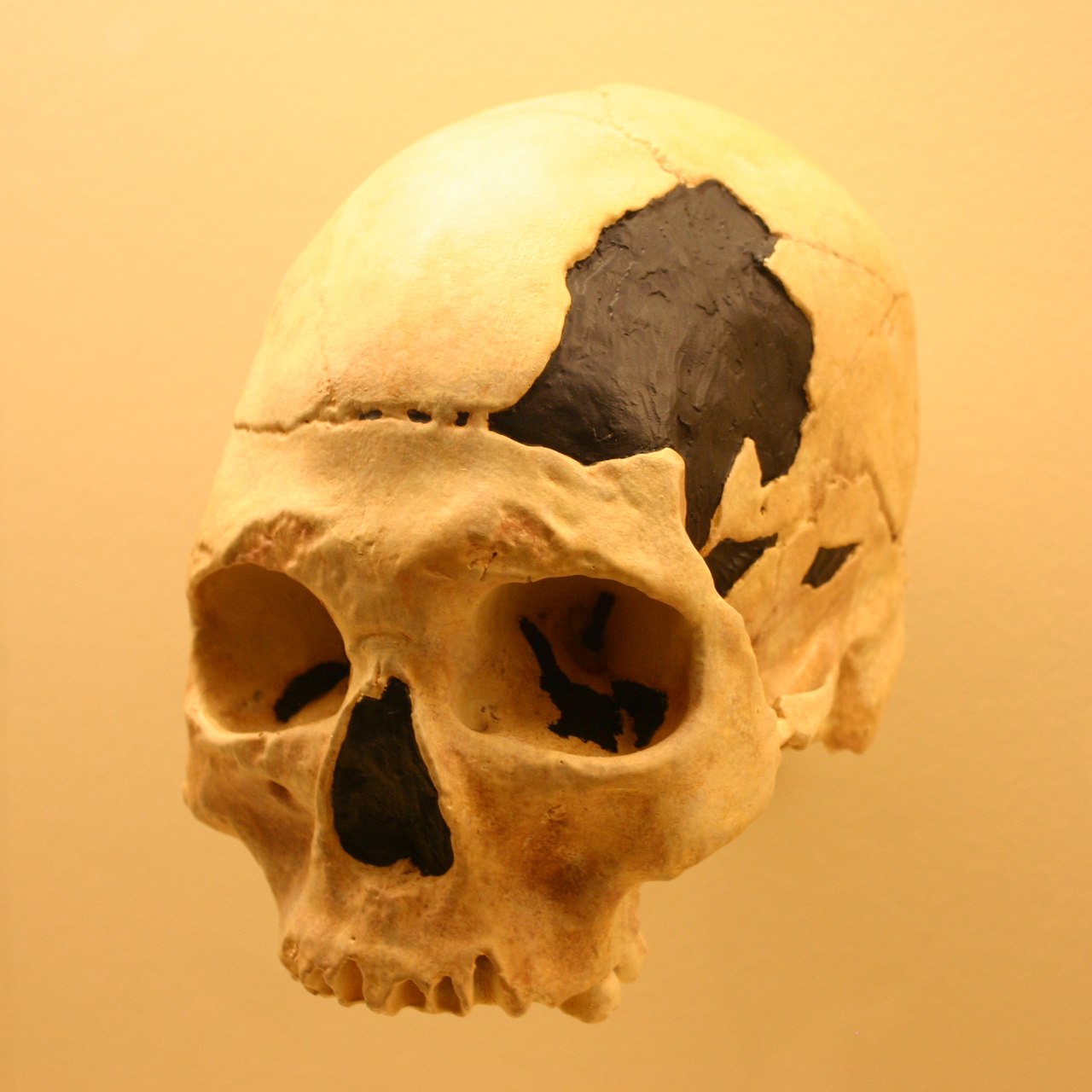 The modern human Oase 2 skull (cast depicted), found in Peştera cu Oase, displays archaic traits due to possible hybridization with Neanderthals.[57] The early modern human Oase 1 mandible from Peștera cu Oase (Romania) of 34,000–36,000 14C years BP presents a mosaic of modern, archaic, and possible Neanderthal features.[58] It displays a lingual bridging of the mandibular foramen, not present in earlier humans except Neanderthals of the late Middle and Late Pleistocene, thus suggesting affinity with Neanderthals.[58] Concluding from the Oase 1 mandible, there was apparently a significant craniofacial change of early modern humans from at least Europe, possibly due to some degree of admixture with Neanderthals.[58] The earliest (before about 33 ka BP) European modern humans and the subsequent (Middle Upper Paleolithic) Gravettians, falling anatomically largely in line with the earliest (Middle Paleolithic) African modern humans, also show traits that are distinctively Neanderthal, suggesting that a solely Middle Paleolithic modern human ancestry was unlikely for European early modern humans.[59] Manot 1, a partial calvarium of a modern human that was recently discovered at the Manot Cave (Western Galilee, Israel) and dated to 54.7±5.5 kyr BP, represents the first fossil evidence from the period when modern humans successfully migrated out of Africa and colonized Eurasia.[60] It also provides the first fossil evidence that modern humans inhabited the southern Levant during the Middle to Upper Palaeolithic interface, contemporaneously with the Neanderthals and close to the probable interbreeding event.[60] The morphological features suggest that the Manot population may be closely related to or may have given rise to the first modern humans who later successfully colonized Europe to establish early Upper Palaeolithic populations.[60] History The interbreeding has been discussed ever since the discovery of Neanderthal remains in the 19th century, though earlier writers believed that Neanderthals were a direct ancestor of modern humans. Thomas Huxley suggested that many Europeans bore traces of Neanderthal ancestry, but associated Neanderthal characteristics with primitivism, writing that since they "belong to a stage in the development of the human species, antecedent to the differentiation of any of the existing races, we may expect to find them in the lowest of these races, all over the world, and in the early stages of all races".[61] Until the early 1950s, most scholars thought Neanderthals were not in the ancestry of living humans.[62][63] Nevertheless, Hans Peder Steensby proposed interbreeding in 1907 in the article Race studies in Denmark. He strongly emphasised that all living humans are of mixed origins.[64] He held that this would best fit observations, and challenged the widespread idea that Neanderthals were ape-like or inferior. Basing his argument primarily on cranial data, he noted that the Danes, like the Frisians and the Dutch, exhibit some Neanderthaloid characteristics, and felt it was reasonable to "assume something was inherited" and that Neanderthals "are among our ancestors". Carleton Stevens Coon in 1962 found it likely, based upon evidence from cranial data and material culture, that Neanderthal and Upper Paleolithic peoples either interbred or that the newcomers reworked Neanderthal implements "into their own kind of tools".[65] By the early 2000s, the majority of scholars supported the Out of Africa hypothesis,[66][67] according to which anatomically modern humans left Africa about 50,000 years ago and replaced Neanderthals with little or no interbreeding. Yet some scholars still argued for hybridisation with Neanderthals. The most vocal proponent of the hybridisation hypothesis was Erik Trinkaus of Washington University in St. Louis.[68] Trinkaus claimed various fossils as products of hybridised populations, including the skeleton of a child found at Lagar Velho in Portugal[69][70][71] and the Peștera Muierii skeletons from Romania.[56] |
形態学 アブリゴ・ド・ラガール・ヴェーリョ(ポルトガル)出土の後期旧石器時代初期の現生人類の子供の埋葬遺物は、ネアンデルタール人がイベリアに分散した現生 人類と交雑したことを示す形質を備えている[55]。埋葬遺物の年代測定(24,500年BP)と、イベリアにおけるネアンデルタール人から現生人類への 移行期(28,000~30,000年BP)以降もネアンデルタール人の形質が長く残っていることを考慮すると、この子供はすでに混血が進んでいた集団の 子孫であった可能性がある[55]。 35,000年BPのペシュテラ・ムイエリロル(ルーマニア)から出土した上旧石器時代初期の現生人類の遺体は、ヨーロッパ現生人類の形態学的パターンを 示しているが、古人あるいはネアンデルタール人の特徴を持っており、ヨーロッパ現生人類がネアンデルタール人と交雑していたことを示唆している[56]。 [56]これらの特徴には、大きな眼窩間幅、比較的平坦な上毛弓、突出した後頭骨饅頭、非対称で浅い下顎ノッチ形状、高い下顎冠状突起、ノッチ稜に対して 相対的に垂直な下顎顆の位置、狭い肩甲骨関節窩が含まれる[56]。 ペシュテラ・ク・オアセで発見された現生人類オアセ2の頭骨(鋳型で描かれている)は、ネアンデルタール人との混血の可能性による古代の特徴を示している[57]。 ルーマニアのPeștera cu Oaseで発見された34,000-36,000年前の初期現生人類Oase 1の下顎骨は、現生人類、古人類、そしてネアンデルタール人の可能性のある特徴のモザイクを示す[58]。 [58] オアーゼ1号の下顎骨から結論すると、少なくともヨーロッパから来た初期の現生人類の頭蓋顔面には、おそらくネアンデルタール人とのある程度の混血による 著しい変化があったようである[58]。 最古の(約33 ka BP以前)ヨーロッパの現生人類とそれに続く(中期上旧石器時代の)グラベット人は、解剖学的には最古の(中期旧石器時代の)アフリカの現生人類とほぼ一 致しているが、ネアンデルタール人特有の形質も示しており、ヨーロッパの現生人類の祖先がもっぱら中期旧石器時代の現生人類である可能性は低いことを示唆 している[59]。 マノット洞窟(イスラエル、西ガリラヤ)で最近発見された、紀元前54.7±5.5kyr BPとされる現生人類の部分的なカルバリウムであるマノット1は、現生人類がアフリカから移動し、ユーラシア大陸を植民地化することに成功した時期の最初 の化石証拠である。 [60] また、この化石は、現生人類が中期から後期旧石器時代の界面において、ネアンデルタール人と同時期に、そして交雑の可能性が高い時期にレバント南部に生息 していたことを示す最初の化石的証拠でもある。 歴史 19世紀にネアンデルタール人の遺骨が発見されて以来、交雑が議論されてきたが、それ以前の作家はネアンデルタール人が現生人類の直接の祖先であると考え ていた。トーマス・ハクスリーは、ヨーロッパ人の多くがネアンデルタール人の祖先の痕跡を持っていることを示唆したが、ネアンデルタール人の特徴を原始主 義と関連付け、「現存するどの種族の分化よりも前の、人類種の発達段階に属するものであるため、世界中のこれらの種族の中で最も下層に位置する種族や、す べての種族の初期段階に見られると予想される」と書いた[61]。 1950年代初頭まで、ほとんどの学者はネアンデルタール人は現生人類の祖先にはいないと考えていた[62][63]。 それにもかかわらず、ハンス・ペーダー・スティーンズビーは1907年にデンマークの人種研究という論文で異種交配を提唱した。彼は、現生人類はすべて混 血起源であることを強く強調した[64]。彼はこれが観察結果に最も合致するとし、ネアンデルタール人は猿に似ている、あるいは劣っているという広範な考 えに異議を唱えた。彼は主に頭蓋のデータに基づいて、デンマーク人がフリジア人やオランダ人と同様にネアンデルタール人の特徴を示していることを指摘し、 「何かが受け継がれたと仮定する」ことは妥当であり、ネアンデルタール人は「我々の祖先の中にいる」と考えた。 1962年のカールトン・スティーブンス・クーンは、頭蓋データと物質文化からの証拠に基づき、ネアンデルタール人と上旧石器時代の人々は交配したか、新参者がネアンデルタール人の道具を「自分たちの種類の道具に作り変えた」可能性が高いと考えた[65]。 2000年代初頭までには、大多数の学者がアウト・オブ・アフリカ仮説を支持していた[66][67]。それによると、解剖学的に現生人類は約5万年前に アフリカを離れ、ネアンデルタール人とほとんど交雑することなく入れ替わったという。それでもなお、ネアンデルタール人との交雑を主張する学者もいた。ト リンカウスは、ポルトガルのラガール・ヴェーリョで発見された子供の骨格[69][70][71]やルーマニアのペシュテラ・ムイエリイの骨格[56]な ど、様々な化石が雑種化した集団の産物であると主張した。 |
| Denisovans Genetics Further information: Australasians § Genetics  The Denisovan genome was sequenced from a fragment of the distal phalanx of the fifth finger (replica depicted) found in the Denisova cave.[23] Proportion of admixture It has been shown that Melanesians (e.g. Papua New Guinean and Bougainville Islander) share relatively more alleles with Denisovans when compared to other Eurasian-derived populations and Africans.[72] It is estimated that 4% to 6% of the genome in Melanesians derives from Denisovans, while no Eurasians or Africans displayed contributions of the Denisovan genes.[72] It has been observed that Denisovans contributed genes to Melanesians but not to East Asians, indicating that there was interaction between the early ancestors of Melanesians with Denisovans but that this interaction did not take place in the regions near southern Siberia, where as-of-yet the only Denisovan remains have been found.[72] In addition, Aboriginal Australians also show a relative increased allele sharing with Denisovans, compared to Eurasians and African populations, consistent with the hypothesis of increased admixture between Denisovans and Melanesians.[73] Reich et al. (2011) produced evidence that the highest presence of Denisovan admixture is in Oceanian populations, followed by many Southeast Asian populations, and none in East Asian populations.[74] There is significant Denisovan genetic material in eastern Southeast Asian and Oceanian populations (e.g. Aboriginal Australians, Near Oceanians, Polynesians, Fijians, eastern Indonesians, Philippine Mamanwa and Manobo), but not in certain western and continental Southeast Asian populations (e.g. western Indonesians, Malaysian Jehai, Andaman Onge, and mainland Asians), indicating that the Denisovan admixture event happened in Southeast Asia itself rather than mainland Eurasia.[74] The observation of high Denisovan admixture in Oceania and the lack thereof in mainland Asia suggests that early modern humans and Denisovans had interbred east of the Wallace Line that divides Southeast Asia according to Cooper and Stringer (2013).[75] Skoglund and Jakobsson (2011) observed that particularly Oceanians, followed by Southeast Asians populations, have a high Denisovans admixture relative to other populations.[76] Furthermore, they found possible low traces of Denisovan admixture in East Asians and no Denisovan admixture in Native Americans.[76] In contrast, Prüfer et al. (2013) found that mainland Asian and Native American populations may have a 0.2% Denisovan contribution, which is about twenty-five times lower than Oceanian populations.[13] The manner of gene flow to these populations remains unknown.[13] However, Wall et al. (2013) stated that they found no evidence for Denisovan admixture in East Asians.[24] Findings indicate that the Denisovan gene flow event happened to the common ancestors of Aboriginal Filipinos, Aboriginal Australians, and New Guineans.[74][77] New Guineans and Australians have similar rates of Denisovan admixture, indicating that interbreeding took place prior to their common ancestors' entry into Sahul (Pleistocene New Guinea and Australia), at least 44,000 years ago.[74] It has also been observed that the fraction of Near Oceanian ancestry in Southeast Asians is proportional to the Denisovan admixture, except in the Philippines where there is a higher proportional Denisovan admixture to Near Oceanian ancestry.[74] Reich et al. (2011) suggested a possible model of an early eastward migration wave of modern humans, some who were Philippine/New Guinean/Australian common ancestors that interbred with Denisovans, respectively followed by divergence of the Philippine early ancestors, interbreeding between the New Guinean and Australian early ancestors with a part of the same early-migration population that did not experience Denisovan gene flow, and interbreeding between the Philippine early ancestors with a part of the population from a much-later eastward migration wave (the other part of the migrating population would become East Asians).[74] Finding components of Denisovan introgression with differing relatedness to the sequenced Denisovan, Browning et al. (2018) suggested that at least two separate episodes of Denisovan admixture has occurred.[78] Specifically, introgression from two distinct Denisovan populations is observed in East Asians (e.g. Japanese and Han Chinese), whereas South Asians (e.g. Telugu and Punjabi) and Oceanians (e.g. Papuans) display introgression from one Denisovan population.[78] Exploring derived alleles from Denisovans, Sankararaman et al. (2016) estimated that the date of Denisovan admixture was 44,000–54,000 years ago.[6] They also determined that the Denisovan admixture was the greatest in Oceanian populations compared to other populations with observed Denisovan ancestry (i.e. America, Central Asia, East Asia, and South Asia).[6] The researchers also made the surprising finding that South Asian populations display an elevated Denisovan admixture (when compared to other non-Oceanian populations with Denisovan ancestry), albeit the highest estimate (which are found in Sherpas) is still ten times lower than in Papuans.[6] They suggest two possible explanations: There was a single Denisovan introgression event that was followed by dilution to different extents or at least three distinct pulses of Denisovan introgressions must have occurred.[6] A study in 2021 analyzing archaic ancestry in 118 Philippine ethnic groups discovered an independent admixture event into Philippine Negritos from Denisovans. The Ayta Magbukon in particular were found to possess the highest level of Denisovan ancestry in the world, with ~30%–40% more than even that found in Australians and Papuans (Australo-Melanesians), suggesting that distinct Islander Denisovan populations existed in the Philippines which admixed with modern humans after their arrival.[79] It has been shown that Eurasians have some but significantly lesser archaic-derived genetic material that overlaps with Denisovans, stemming from the fact that Denisovans are related to Neanderthals—who contributed to the Eurasian gene pool—rather than from interbreeding of Denisovans with the early ancestors of those Eurasians.[23][72] The skeletal remains of an early modern human from the Tianyuan cave (near Zhoukoudian, China) of 40,000 years BP showed a Neanderthal contribution within the range of today's Eurasian modern humans, but it had no discernible Denisovan contribution.[80] It is a distant relative to the ancestors of many Asian and Native American populations, but post-dated the divergence between Asians and Europeans.[80] The lack of a Denisovan component in the Tianyuan individual suggests that the genetic contribution had been always scarce in the mainland.[13] Reduced contribution There are large genomic regions devoid of Denisovan-derived ancestry, partly explained by infertility of male hybrids, as suggested by the lower proportion of Denisovan-derived ancestry on X chromosomes and in genes that are expressed in the testes of modern humans.[6] Changes in modern humans Exploring the immune system's HLA alleles, it has been suggested that HLA-B*73 introgressed from Denisovans into modern humans in western Asia due to the distribution pattern and divergence of HLA-B*73 from other HLA alleles.[48] Even though HLA-B*73 is not present in the sequenced Denisovan genome, HLA-B*73 was shown to be closely associated to the Denisovan-derived HLA-C*15:05 from the linkage disequilibrium.[48] From phylogenetic analysis, however, it has been concluded that it is highly likely that HLA-B*73 was ancestral.[44] The Denisovan's two HLA-A (A*02 and A*11) and two HLA-C (C*15 and C*12:02) allotypes correspond to common alleles in modern humans, whereas one of the Denisovan's HLA-B allotype corresponds to a rare recombinant allele and the other is absent in modern humans.[48] It is thought that these must have been contributed from Denisovans to modern humans, because it is unlikely to have been preserved independently in both for so long due to HLA alleles' high mutation rate.[48] Tibetan people received an advantageous EGLN1 and EPAS1 gene variant, associated with hemoglobin concentration and response to hypoxia, for life at high altitudes from the Denisovans.[44] The ancestral variant of EPAS1 upregulates hemoglobin levels to compensate for low oxygen levels—such as at high altitudes—but this also has the maladaption of increasing blood viscosity.[81] The Denisovan-derived variant on the other hand limits this increase of hemoglobin levels, thus resulting in a better altitude adaption.[81] The Denisovan-derived EPAS1 gene variant is common in Tibetans and was positively selected in their ancestors after they colonized the Tibetan plateau.[81] |
デニソワ人 遺伝学 さらに詳しい情報 オーストラリア人§遺伝学 デニソワ人のゲノムはデニソワ洞窟で発見された第5指の遠位指骨の断片(レプリカ)から解読された[23]。 混血の割合 メラネシア人(パプアニューギニア人やブーゲンビル島民など)は、他のユーラシア由来の集団やアフリカ人と比較して、デニソワ人と比較的多くの対立遺伝子 を共有していることが示されている[72]。メラネシア人のゲノムの4%から6%がデニソワ人に由来すると推定されているが、ユーラシア人やアフリカ人に はデニソワ人の遺伝子の寄与は見られない。 このことは、メラネシア人の初期の祖先とデニソワ人との間に相互作用があったことを示しているが、この相互作用は、まだ唯一のデニソワ人の遺骨が発見され ている南シベリア付近の地域では起こらなかったことを示している[72]。 [さらに、オーストラリアのアボリジニも、ユーラシア人やアフリカの集団と比較して、デニソワ人との対立遺伝子の共有が相対的に増加しており、デニソワ人 とメラネシア人との混血が増加したという仮説と一致している[73]。 Reichら(2011)は、デニソワ人の混血が最も多いのはオセアニアの集団で、次いで多くの東南アジアの集団であり、東アジアの集団では皆無であると いう証拠を示している[74]。東南アジアとオセアニアの東部集団(例:オーストラリア原住民、近オセアニア人、ポリネシア人、フィジー人、インドネシア 東部、フィリピンのママンワ人、マノボ人)にはかなりのデニソワ人の遺伝物質が存在するが、東南アジアの西部と大陸の特定の集団(例:インドネシア西部、 マレーシア人)には存在しない。 74]。オセアニアでデニソワ人との混血が多く、アジア本土では見られないことから、クーパーとストリンガー(2013年)によれば、初期現生人類とデニ ソワ人は東南アジアを分断するウォレス線の東側で交雑していたことが示唆されている[75]。 SkoglundとJakobsson(2011)は、特にオセアニア人、次いで東南アジア人の集団が他の集団に比べてデニソワ人との混血が多いことを観 察している[76]。さらに彼らは、東アジア人ではデニソワ人との混血の痕跡が低い可能性があり、アメリカ先住民ではデニソワ人との混血がないことを発見 している[76]。(2013)は、アジア本土とネイティブアメリカンの集団は0.2%のデニソワの寄与がある可能性があり、これはオセアニアの集団より も約25倍低いことを明らかにした[13]。 フィリピンのアボリジニ、オーストラリアのアボリジニ、ニューギニア人の共通の祖先にデニソワ人の遺伝子流入が起こったことを示す調査結果がある[74] [77]。ニューギニア人とオーストラリア人はデニソワ人の混血率がほぼ同じであり、少なくとも44,000年前に共通の祖先がサフル(更新世ニューギニ アとオーストラリア)に入る前に交雑が起こったことを示している。 [74]また、東南アジア人の近オセアニア系祖先の割合はデニソワ人との混血に比例しているが、フィリピンでは近オセアニア系祖先に対するデニソワ人との 混血の割合が高い。 [74] Reich et al. (2011)は、フィリピン/ニューギニア/オーストラリアの共通祖先がそれぞれデニソワ人と交雑し、その後フィリピンの初期祖先が分岐したという、現生 人類の初期の東方への移動の波のモデルの可能性を示唆した、 ニューギニア人とオーストラリア人の初期祖先と、デニソワ人の遺伝子流を経験しなかった同じ初期移住集団の一部との交雑、フィリピンの初期祖先と、それよ りずっと後の東方移住の波の集団の一部との交雑(移住集団の他の一部は東アジア人になる)。 [74] 塩基配列を決定したデニソワ人との関連性が異なるデニソワ人の内進の構成要素を発見したブラウニングら(2018年)は、デニソワ人の混血の少なくとも2 つの別々のエピソードが発生したことを示唆した。 [78]具体的には、東アジア人(日本人や漢民族など)では2つの異なるデニソワ集団からの導入が観察されるのに対し、南アジア人(テルグ人やパンジャブ 人など)やオセアニア人(パプア人など)では1つのデニソワ集団からの導入が見られる[78]。 Sankararamanら(2016年)は、デニソワ人からの派生対立遺伝子を探索し、デニソワ人の混血の年代は44,000-54,000年前である と推定した[6]。彼らはまた、デニソワ人の混血は、デニソワ人の祖先が観察された他の集団(すなわち、アメリカ、中央アジア、東アジア、南アジア)と比 較して、オセアニア人の集団で最大であると決定した。 [6]研究者たちはまた、(デニソワ人の祖先を持つ他の非オセアニア系集団と比較した場合)南アジア系集団がデニソワ人との混血が高いという驚くべき発見 をした: すなわち、単一のデニソワ人導入事象があり、その後に異なる程度まで希釈されたか、あるいは少なくとも3つの異なるデニソワ人導入のパルスが発生したに違 いないということである[6]。 2021年に行われたフィリピンの118の民族グループの古代の祖先を分析した研究では、デニソワ人からフィリピンのネグリト人への独立した混血事象が発 見された。特にアヤタ・マグブコンは、オーストラリア人やパプア人(アウストラロ・メラネシア人)よりも30%~40%多い、世界で最も高いレベルのデニ ソワ人の祖先を持っていることが発見され、フィリピンに別個の島民デニソワ人の集団が存在し、彼らが到着した後に現生人類と混血したことが示唆された [79]。 ユーラシア人はデニソワ人と重なる古人由来の遺伝物質をいくつか持っているが、かなり少ないことが示されており、これはデニソワ人がユーラシア人の初期の 祖先と交雑したためというよりも、デニソワ人がユーラシア人の遺伝子プールに貢献したネアンデルタール人と近縁であるという事実に由来している[23] [72]。 4万年前の天元洞窟(中国の周口店の近く)から出土した初期現生人類の骨格は、今日のユーラシア現生人類の範囲内でネアンデルタール人の寄与を示したが、 デニソワ人の寄与は識別できなかった。 [80]この個体は多くのアジア人やアメリカ先住民の祖先の遠い親戚であるが、アジア人とヨーロッパ人の分岐より後のものであった[80]。天元の個体に デニソワ人の成分がないことは、遺伝的な寄与が本土では常に希薄であったことを示唆している[13]。 寄与の減少 現代人のX染色体や精巣で発現する遺伝子におけるデニソワ人由来の祖先の割合が低いことから示唆されるように、デニソワ人由来の祖先が存在しないゲノム領域が広く存在する。 現代人の変化 免疫系のHLA対立遺伝子を探索すると、HLA-B*73の分布パターンと他のHLA対立遺伝子との分岐から、HLA-B*73がデニソワ人から西アジア の現生人類に導入されたことが示唆されている。 [48]HLA-B*73は解読されたデニソワ人のゲノムには存在しないにもかかわらず、連鎖不平衡からHLA-B*73はデニソワ人由来のHLA- C*15:05と密接に関連していることが示された[48]。しかし系統学的解析から、HLA-B*73が祖先的であった可能性が高いと結論されている [44]。 デニソワ人の2つのHLA-Aアロタイプ(A*02とA*11)と2つのHLA-Cアロタイプ(C*15とC*12:02)は現生人類の共通対立遺伝子に 対応し、デニソワ人のHLA-Bアロタイプの1つは稀な組換え対立遺伝子に対応し、もう1つは現生人類には存在しない。 [48]。HLA対立遺伝子は変異率が高いため、長い間両者で独立して保存されていたとは考えにくいため、これらはデニソワ人から現生人類に寄与したに違 いないと考えられている[48]。 チベット人は、高地での生活に有利なヘモグロビン濃度と低酸素への反応に関連するEGLN1とEPAS1遺伝子の変種をデニソワ人から受け継いだ [44]。EPAS1の祖先の変種は、高地などの低酸素レベルを補うためにヘモグロビン濃度を上昇させるが、これには血液粘度を増加させるという不適応も ある。 [一方、デニソワ人由来の変異型は、このヘモグロビン濃度の上昇を制限するため、より優れた高度適応をもたらす[81]。デニソワ人由来のEPAS1遺伝 子の変異型はチベット人に多く、彼らがチベット高原を植民地化した後、彼らの祖先において正選択された[81]。 |
| Archaic African hominins Rapid decay of fossils in Sub-Saharan African environments makes it currently unfeasible to compare modern human admixture with reference samples of archaic Sub-Saharan African hominins.[5][82] Ancient DNA Data from a ~4,500 BP Ethiopian highland individual,[83] and from Southern (~2,300–1,300 BP), and Eastern and South-Central Africa (~8,100–400 BP) has clarified that some West Africa populations have small amounts of excess alleles best explained by an archaic source in West Africans that is not included in the pre-agricultural Eastern African hunter-gatherers, Southern African hunter-gatherer populations, or the genetic gradation between them. The West African groups carrying the archaic DNA include Yoruba from coastal Nigeria and Mende from Sierra Leon indicating that the ancient DNA was acquired long before the spread of agriculture and likely well before the Holocene (starting 11,600 BP), Such an archaic lineage must have separated before the divergence of San ancestors, which is estimated to have begun on the order of 200–300 thousand years ago.[84][85] The hypothesis that there has been archaic line in the ancestry of present-day Africans that originated before the San, Pygmies and East African hunter gatherers (and the Eurasians) is supported by a line of evidence independent from the Skoglund findings based on long haplotypes with deep divergences from other human haplotypes including Lachance et al.(2012),[82] Hammer et al., 2011,[86] and Plagnol and Wall (2006).[87] In the archaic DNA differences found by Hammer, et al., the pygmies (of Central Africa) are grouped with the San (of Southern Africa) in contrast to the Yoruba (of West Africa). Further clarification of the presence of archaic DNA in current West African populations with the extraction and sequencing of DNA from 4 fossils found at Shum Laka in Cameroon dating from 8,000 to 3,000 BP. These individuals were found to derive most of their DNA from Central African hunter gatherers (Pygmy ancestors) and did not share the archaic DNA found in the Yoruba and Mande.[88] The pattern of differences between Eastern, Central and Southern hunter gatherers when compared to the West African groups which had been found by Hammer was confirmed. In a second study Lipson et al. (2020) studied DNA extracted from 6 additional Eastern and Southcentral African fossils from the last 18,000 years. It was determined that their genetic origins could be accounted for by DNA contributions from Southern, Central and Eastern hunter gatherers, and that none of them had the archaic DNA found in the Yoruba.[89] According to a study published in 2020, there are indications that 2% to 19% (or about ≃6.6 and ≃7.0%) of the DNA of four West African populations may have come from an unknown archaic hominin which split from the ancestor of humans and Neanderthals between 360 kya to 1.02 mya. However, in contrast to the studies of Skoglund and Lipson with ancient African DNA, the study also finds that at least part of this proposed archaic admixture is also present in Eurasians/non-Africans, and that the admixture event or events range from 0 to 124 ka B.P, which includes the period before the Out-of-Africa migration and prior to the African/Eurasian split (thus affecting in part the common ancestors of both Africans and Eurasians/non-Africans).[90][91][92] Another recent study, which discovered substantial amounts of previously undescribed human genetic variation, also found ancestral genetic variation in Africans that predates modern humans and was lost in most non-Africans.[8] |
古代のアフリカ原人 サハラ以南のアフリカの環境における化石の急速な腐敗によって、古代のサハラ以南のアフリカのホミニンの参照サンプルと現生人類の混血を比較することは現在のところ不可能である[5][82]。 約4,500BPのエチオピア高地の個体[83]、南部アフリカ(~2,300~1,300BP)、東部・南中部アフリカ(~8,100~400BP)の 古代DNAデータから、西アフリカの集団の中には、農業以前の東部アフリカの狩猟採集民、南部アフリカの狩猟採集民、またはそれらの間の遺伝的グラデー ションには含まれない、西アフリカ人の古代のソースによって最もよく説明される少量の過剰対立遺伝子を持つ集団があることが明らかになった。古代のDNA を持つ西アフリカのグループには、ナイジェリア沿岸部のヨルバやシエラレオンのメンデが含まれ、古代のDNAは農耕が広まるずっと前、完新世 (11,600BPから始まる)よりかなり前に獲得された可能性が高いことを示している。 現在のアフリカ人の祖先には、サン、ピグミー、東アフリカの狩猟採集民(およびユーラシア人)以前に起源を持つ古代の系統が存在するという仮説は、 Lachanceら(2012年)[82]、Hammerら(2011年)[86]、PlagnolとWall(2006年)[87]を含む他のヒトのハ プロタイプと深い分岐を持つ長いハプロタイプに基づくスコグランド所見とは独立した一連の証拠によって支持されている。 Hammerらによって発見された古代のDNAの違いでは、ピグミー(中央アフリカ)はヨルバ(西アフリカ)とは対照的にサン(南部アフリカ)とグループ 化されている。カメルーンのシュム・ラカで発見された紀元前8,000年から3,000年前の4つの化石からDNAを抽出し、塩基配列を決定したことで、 現在の西アフリカの集団に古代のDNAが存在することがさらに明らかになった。これらの個体は、そのDNAのほとんどが中央アフリカの狩猟採集民(ピグ ミーの祖先)に由来し、ヨルバやマンデに見られる古代のDNAを共有していないことが判明した[88]。ハマーによって発見された西アフリカの集団と比較 した場合、東部、中部、南部の狩猟採集民の間に違いがあるというパターンが確認された。リプソンら(2020年)は2つ目の研究で、過去18,000年間 に産出したアフリカ東部と中南部の化石6点から抽出したDNAを調査した。その結果、彼らの遺伝的起源は南部、中部、東部の狩猟採集民からのDNAの寄与 で説明でき、ヨルバ族に見られるような古代のDNAを持つものはいなかったことが判明した[89]。 2020年に発表された研究によると、西アフリカの4つの集団のDNAの2%から19%(または約6.6%と約7.0%)が、360kyaから 1.02myaの間にヒトとネアンデルタール人の祖先から分裂した未知の古人類に由来する可能性が指摘されている。しかし、Skoglundと Lipsonによる古代アフリカのDNAを用いた研究とは対照的に、本研究では、この古代の混血の少なくとも一部はユーラシア人/非アフリカ人にも存在 し、その混血は紀元前0年から124kaの範囲に及ぶことも発見している。 また、この混血は、アフリカ外からの移住以前、アフリカ人とユーラシア人の分裂以前の期間を含む(したがって、アフリカ人とユーラシア人/非アフリカ人の 共通の祖先に一部影響を与えている)[90][91][92]。また、別の最近の研究では、これまで記述されていなかったヒトの遺伝的変異が相当量発見さ れ、アフリカ人の祖先の遺伝的変異は、現生人類より前のものであり、ほとんどの非アフリカ人では失われていることも発見された[8]。 |
| Archaic hominins in Eurasia Hominins' presence in Eurasia begun at least 2 million years BP. Genetic evidence shows that thousands of years later when lineages of Neandertals and Denisovans started to expand into Eurasia, the continent was still inhabited by descendants of these archaic hominins, and their genetic admixture made its way into genome of Neanderthals and Denisovans and later indirectly into modern humans.[93][94] Genetic studies show two major events of genetic admixture from superarchaics, suggesting that in the late middle Pleistocene, Eurasia was inhabited by at least two separate populations of ancient hominins.[93] Roger et al. (2020) describes an event of admixture that occurred soon after Neandersovans (common ancestor of Neanderthals and Denisovans) started to expand into Eurasia. They met a lineage of superarchaic hominins that had been separated from African homo lineages since at least 2 Ma ago.[93] Previous studies identified more recent event of admixture. About 350,000 years ago a genome of an "erectus-like" creature was injected into the Denisovan lineage. With the separation time of about 2 Ma ago and interbreeding that happened 350 ka ago, the two populations involved were more distantly related than any pair of human populations previously known to interbreed.[93][94] |
ユーラシア大陸の古人類 ユーラシア大陸におけるヒトの存在は、少なくとも200万年前に始まった。数千年後にネアンデルタール人とデニソワ人の系統がユーラシア大陸に進出し始め たとき、大陸にはまだこれらの古人類の子孫が住んでおり、彼らの遺伝的混血がネアンデルタール人とデニソワ人のゲノムに入り込み、後に間接的に現生人類に 入り込んだことが遺伝学的証拠によって示されている[93][94]。 遺伝学的研究は、超古人類からの遺伝的混血の2つの主要な出来事を示しており、更新世後期のユーラシア大陸には少なくとも2つの別個の古代ヒトの集団が居住していたことを示唆している[93]。 Rogerら(2020年)は、ネアンデルソヴァ人(ネアンデルタール人とデニソワ人の共通祖先)がユーラシア大陸に進出し始めた直後に起こった混血の出 来事について述べている。彼らは、少なくとも2Ma前以降アフリカのホモ系統から分離していた超古代ホミニンの系統に出会ったのである[93]。 以前の研究では、混血はもっと最近の出来事であることが確認されている。約35万年前、「エレクタスに似た」生物のゲノムがデニソワ人の系統に注入され た。約2Ma前の分離時期と350Ka前に起こった交雑により、関与した2つの集団は、これまで交雑が知られていたどのヒト集団のペアよりも遠縁であった [93][94]。 |
| Related studies In 2019, scientists discovered evidence, based on genetics studies using artificial intelligence (AI), that suggests the existence of an unknown human ancestor species, not Neanderthal or Denisovan, in the genome of modern humans.[95][96] |
関連研究 2019年、科学者たちは人工知能(AI)を用いた遺伝学的研究に基づき、ネアンデルタール人でもデニソワ人でもない未知の人類の祖先種が現生人類のゲノムに存在することを示唆する証拠を発見した[95][96]。 |
| Interbreeding Multiregional origin of modern humans Neanderthal extinction |
異種交配 現生人類の多地域的起源 ネアンデルタール人の絶滅 |
| https://en.wikipedia.org/wiki/Interbreeding_between_archaic_and_modern_humans |
リ ンク
文 献
そ の他の情報
Copyleft, CC, Mitzub'ixi Quq Chi'j, 1996-2099
![]()
☆
 ☆
☆