
音楽の神経科学
Neuroscience of music
Francis Miller American, 1905–1973 Listening to music in art class, 1957
☆ 音楽の神経科学とは、音楽の認知プロセスに関わる脳のメカニズムを科学的に研究する学問である。こうした行動には、音楽鑑賞、演奏、作曲、読譜、作曲、付 随する活動などが含まれる。また、音楽美学や音楽感情の脳内メカニズムについても、ますます関心が高まっている。この分野の研究者は、認知神経科学、神経 学、神経解剖学、心理学、音楽理論、コンピュータサイエンス、その他の関連分野のトレーニングを受けている場合がある。 音楽認知神経科学は音楽心理学の重要な一分野であり、脳の直接観察と機能的磁気共鳴画像法(fMRI)や陽電子放射断層撮影(PET)などの脳画像技術の 使用に重点を置いている点で、認知音楽学などの関連分野とは区別される。
| The neuroscience of
music is the scientific study of brain-based mechanisms involved in the
cognitive processes underlying music. These behaviours include music
listening, performing, composing, reading, writing, and ancillary
activities. It also is increasingly concerned with the brain basis for
musical aesthetics and musical emotion. Scientists working in this
field may have training in cognitive neuroscience, neurology,
neuroanatomy, psychology, music theory, computer science, and other
relevant fields. The cognitive neuroscience of music represents a significant branch of music psychology, and is distinguished from related fields such as cognitive musicology in its reliance on direct observations of the brain and use of brain imaging techniques like functional magnetic resonance imaging (fMRI) and positron emission tomography (PET). |
音楽の神経科学とは、音楽の認知プロセスに関わる脳のメカニズムを科学
的に研究する学問である。こうした行動には、音楽鑑賞、演奏、作曲、読譜、作曲、付随する活動などが含まれる。また、音楽美学や音楽感情の脳内メカニズム
についても、ますます関心が高まっている。この分野の研究者は、認知神経科学、神経学、神経解剖学、心理学、音楽理論、コンピュータサイエンス、その他の
関連分野のトレーニングを受けている場合がある。 音楽認知神経科学は音楽心理学の重要な一分野であり、脳の直接観察と機能的磁気共鳴画像法(fMRI)や陽電子放射断層撮影(PET)などの脳画像技術の 使用に重点を置いている点で、認知音楽学などの関連分野とは区別される。 |
| Elements of music Pitch Sounds consist of waves of air molecules that vibrate at different frequencies. These waves travel to the basilar membrane in the cochlea of the inner ear. Different frequencies of sound will cause vibrations in different locations of the basilar membrane. We are able to hear different pitches because each sound wave with a unique frequency is correlated to a different location along the basilar membrane. This spatial arrangement of sounds and their respective frequencies being processed in the basilar membrane is known as tonotopy. When the hair cells on the basilar membrane move back and forth due to the vibrating sound waves, they release neurotransmitters and cause action potentials to occur down the auditory nerve. The auditory nerve then leads to several layers of synapses at numerous clusters of neurons, or nuclei, in the auditory brainstem. These nuclei are also tonotopically organized, and the process of achieving this tonotopy after the cochlea is not yet well understood.[1] This tonotopy is in general maintained up to primary auditory cortex in mammals.[2] A widely postulated mechanism for pitch processing in the early central auditory system is the phase-locking and mode-locking of action potentials to frequencies in a stimulus. Phase-locking to stimulus frequencies has been shown in the auditory nerve,[3][4] the cochlear nucleus,[3][5] the inferior colliculus,[6] and the auditory thalamus.[7] By phase- and mode-locking in this way, the auditory brainstem is known to preserve a good deal of the temporal and low-passed frequency information from the original sound; this is evident by measuring the auditory brainstem response using EEG.[8] This temporal preservation is one way to argue directly for the temporal theory of pitch perception, and to argue indirectly against the place theory of pitch perception. 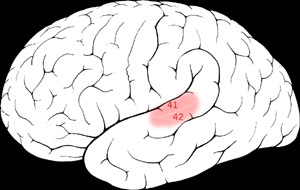 The primary auditory cortex is one of the main areas associated with superior pitch resolution. The right secondary auditory cortex has finer pitch resolution than the left. Hyde, Peretz and Zatorre (2008) used functional magnetic resonance imaging (fMRI) in their study to test the involvement of right and left auditory cortical regions in the frequency processing of melodic sequences.[9] As well as finding superior pitch resolution in the right secondary auditory cortex, specific areas found to be involved were the planum temporale (PT) in the secondary auditory cortex, and the primary auditory cortex in the medial section of Heschl's gyrus (HG). Many neuroimaging studies have found evidence of the importance of right secondary auditory regions in aspects of musical pitch processing, such as melody.[10] Many of these studies such as one by Patterson, Uppenkamp, Johnsrude and Griffiths (2002) also find evidence of a hierarchy of pitch processing. Patterson et al. (2002) used spectrally matched sounds which produced: no pitch, fixed pitch or melody in an fMRI study and found that all conditions activated HG and PT. Sounds with pitch activated more of these regions than sounds without. When a melody was produced activation spread to the superior temporal gyrus (STG) and planum polare (PP). These results support the existence of a pitch processing hierarchy. |
音楽の要素 音高 音は、異なる周波数で振動する空気分子の波で構成されている。これらの波は、内耳の蝸牛にある基底膜に伝わる。異なる周波数の音は、基底膜の異なる位置で 振動を引き起こす。異なる音の高さを聞き分けられるのは、固有の周波数を持つそれぞれの音波が、基底膜の異なる位置と相関しているためである。この音の空 間配置と、基底膜で処理されるそれぞれの周波数は、トノトピーとして知られている。音波の振動により基底膜の有毛細胞が前後に動くと、神経伝達物質が放出 され、聴神経に活動電位が発生する。聴神経は、聴覚脳幹の多数のニューロン群または核のシナプス層へとつながっている。これらの核もまた音高順に組織化さ れており、蝸牛の後にこの音高順性がどのようにして達成されるのかは、まだ十分に解明されていない。[1] この音高順性は、一般的に哺乳類の一次聴覚野まで維持されている。[2] 初期の中枢聴覚系における音高処理のメカニズムとして広く仮定されているのは、刺激の周波数に対する活動電位の位相同期とモード同期である。刺激の周波数 に対する位相同期は、聴神経[3][4]、蝸牛神経核[3][5]、下丘[6]、聴覚野[7]で確認されている。 このように位相およびモードの固定によって、聴性脳幹は元の音からかなりの量の時間的およびローパス周波数情報を保持することが知られている。これは、 EEG(脳波)を使用して聴性脳幹反応を測定することで明らかである。[8] この時間的保存は、時間理論による音高知覚を直接的に主張し、間接的に場所理論による音高知覚を否定する一つの方法である。 一次聴覚野は、優れた音程分解能に関連する主な領域のひとつである。 右の二次聴覚野は左よりも音程分解能が高い。Hyde、Peretz、Zatorre(2008年)は、旋律のシーケンスの周波数処理における左右の聴覚 皮質領域の関与を検証するために、機能的磁気共鳴画像法(fMRI)を用いた研究を行った。 [9] その結果、右側二次聴覚野で優れた音程分解能が認められたほか、特定の領域が関与していることが判明した。その領域とは、二次聴覚野の側頭葉平面(PT) と、ヘシュル回(HG)の内側部分にある一次聴覚野である。 多くの神経画像研究が、メロディなどの音楽の音程処理の側面において、右側二次聴覚領域が重要な役割を果たしているという証拠を発見している。[10] パターソン、ウッペンカンプ、ジョンズルーデ、グリフィス(2002年)による研究など、これらの研究の多くは、音程処理の階層構造の証拠も発見してい る。Patterson ら(2002)は、fMRI研究において、スペクトルが一致した音を使用し、ピッチなし、固定ピッチ、メロディを生成した。その結果、すべての条件でHG とPTが活性化することが分かった。ピッチのある音は、ピッチのない音よりもこれらの領域をより活性化した。メロディが生成されると、活性化は上側頭回 (STG)と極平面(PP)に広がった。これらの結果は、ピッチ処理の階層構造の存在を裏付けるものである。 |
Absolute pitch Musicians possessing perfect pitch can identify the pitch of musical tones without external reference. Main article: Absolute pitch Absolute pitch (AP) is defined as the ability to identify the pitch of a musical tone or to produce a musical tone at a given pitch without the use of an external reference pitch.[11][12] Neuroscientific research has not discovered a distinct activation pattern common for possessors of AP. Zatorre, Perry, Beckett, Westbury and Evans (1998) examined the neural foundations of AP using functional and structural brain imaging techniques.[13] Positron emission tomography (PET) was utilized to measure cerebral blood flow (CBF) in musicians possessing AP and musicians lacking AP. When presented with musical tones, similar patterns of increased CBF in auditory cortical areas emerged in both groups. AP possessors and non-AP subjects demonstrated similar patterns of left dorsolateral frontal activity when they performed relative pitch judgments. However, in non-AP subjects activation in the right inferior frontal cortex was present whereas AP possessors showed no such activity. This finding suggests that musicians with AP do not need access to working memory devices for such tasks. These findings imply that there is no specific regional activation pattern unique to AP. Rather, the availability of specific processing mechanisms and task demands determine the recruited neural areas. |
絶対音感 絶対音感を持つ音楽家は、外部参照なしに音の高さを特定することができる。 詳細は「絶対音感」を参照 絶対音感(AP)とは、外部参照音高を使用せずに、音の高さを特定したり、特定の高さの音を出す能力と定義される。[11][12] 神経科学の研究では、絶対音感を持つ人々に共通する明確な活性化パターンは発見されていない。Zatorre、Perry、Beckett、 Westbury、Evans(1998年)は、機能的および構造的脳画像技術を用いて、APの神経基盤を調査した。[13] 陽電子放射断層撮影(PET)を用いて、APを持つ音楽家と持たない音楽家の脳血流(CBF)を測定した。音楽音を提示したところ、両グループで聴覚皮質 領域における同様のCBF増加パターンが現れた。AP保有者と非保有者は、相対音高の判断を行う際の左背外側前頭前野の活動パターンが類似していた。しか し、非保有者では右下前頭前野の活性化が見られたのに対し、AP保有者ではそのような活性化は見られなかった。この発見は、AP保有者の音楽家は、そのよ うな作業を行う際にワーキングメモリーにアクセスする必要がないことを示唆している。これらの発見は、AP保有者に特有の特定の活性化パターンは存在しな いことを示唆している。むしろ、特定の処理メカニズムの有効性とタスクの要求が、動員される神経領域を決定する。 |
| Melody Studies suggest that individuals are capable of automatically detecting a difference or anomaly in a melody such as an out of tune pitch which does not fit with their previous music experience. This automatic processing occurs in the secondary auditory cortex. Brattico, Tervaniemi, Naatanen, and Peretz (2006) performed one such study to determine if the detection of tones that do not fit an individual's expectations can occur automatically.[14] They recorded event-related potentials (ERPs) in nonmusicians as they were presented unfamiliar melodies with either an out of tune pitch or an out of key pitch while participants were either distracted from the sounds or attending to the melody. Both conditions revealed an early frontal error-related negativity independent of where attention was directed. This negativity originated in the auditory cortex, more precisely in the supratemporal lobe (which corresponds with the secondary auditory cortex) with greater activity from the right hemisphere. The negativity response was larger for pitch that was out of tune than that which was out of key. Ratings of musical incongruity were higher for out of tune pitch melodies than for out of key pitch. In the focused attention condition, out of key and out of tune pitches produced late parietal positivity. The findings of Brattico et al. (2006) suggest that there is automatic and rapid processing of melodic properties in the secondary auditory cortex.[14] The findings that pitch incongruities were detected automatically, even in processing unfamiliar melodies, suggests that there is an automatic comparison of incoming information with long term knowledge of musical scale properties, such as culturally influenced rules of musical properties (common chord progressions, scale patterns, etc.) and individual expectations of how the melody should proceed. |
旋律 研究によると、個人は、これまでの音楽経験に合わない音程のずれなど、旋律における相違や異常を自動的に検出できることが示唆されている。この自動処理 は、二次聴覚野で行われる。Brattico、Tervaniemi、Naatanen、Peretz(2006年)は、個人の期待に合わない音程を自動 的に検出できるかどうかを調べる研究を行った。[14] 彼らは、音から注意をそらしている場合とメロディに注意を向けている場合の両方で、音程が合っていない音やキーが合っていない音を提示し、非音楽家の事象 関連電位(ERP)を記録した。両方の条件において、注意が向けられた場所に関わらず、早期の前頭葉エラー関連陰性電位が明らかになった。この陰性電位は 聴覚野、より正確には側頭葉上(これは二次聴覚野に対応する)で発生し、右半球の活動がより活発であった。音程が外れた音に対する陰性電位反応は、音程が 外れた音に対する反応よりも大きかった。音楽的な不調和の評価は、音程が外れたメロディの方が音高が外れたメロディよりも高かった。注意を集中させた条件 では、音高が外れた音程と音高が外れた音程は、後頭部の陽性反応を引き起こした。Brattico らの研究結果(2006年)は、 (2006年)の研究結果は、二次聴覚野において旋律の特性が自動的かつ迅速に処理されていることを示唆している。[14] ピッチの不調和が、たとえ聞き慣れない旋律の処理においても自動的に検出されるという結果は、入力情報と音楽の音階特性に関する長期的知識(音楽的特性に 関する文化の影響を受けた規則(一般的なコード進行、音階パターンなど)や、旋律がどのように進行すべきかに関する個人の期待など)が自動的に比較されて いることを示唆している。 |
| Rhythm The belt and parabelt areas of the right hemisphere are involved in processing rhythm.[15] Rhythm is a strong repeated pattern of movement or sound. When individuals are preparing to tap out a rhythm of regular intervals (1:2 or 1:3) the left frontal cortex, left parietal cortex, and right cerebellum are all activated. With more difficult rhythms such as a 1:2.5, more areas in the cerebral cortex and cerebellum are involved.[16] EEG recordings have also shown a relationship between brain electrical activity and rhythm perception. Snyder and Large (2005)[17] performed a study examining rhythm perception in human subjects, finding that activity in the gamma band (20 – 60 Hz) corresponds to the beats in a simple rhythm. Two types of gamma activity were found by Snyder & Large: induced gamma activity, and evoked gamma activity. Evoked gamma activity was found after the onset of each tone in the rhythm; this activity was found to be phase-locked (peaks and troughs were directly related to the exact onset of the tone) and did not appear when a gap (missed beat) was present in the rhythm. Induced gamma activity, which was not found to be phase-locked, was also found to correspond with each beat. However, induced gamma activity did not subside when a gap was present in the rhythm, indicating that induced gamma activity may possibly serve as a sort of internal metronome independent of auditory input. |
リズム 右半球のベルトおよびパラベルト領域は、リズムの処理に関与している。[15] リズムとは、動きまたは音の強い反復パターンである。個人が規則的な間隔(1:2または1:3)のリズムをタップする準備をしている場合、左前頭皮質、左 頭頂皮質、および右小脳がすべて活性化される。1:2.5 などのより複雑なリズムでは、大脳皮質と小脳のより広範囲の領域が関与する。[16] また、脳波の記録からも、脳の電気的活動とリズム知覚の関係が示されている。 Snyder と Large (2005)[17] は、被験者のリズム知覚を調査する研究を実施し、単純なリズムのビートにガンマ帯域(20 – 60 Hz)の活動が対応していることを発見した。Snyder & Large によって、誘導ガンマ活動と誘発ガンマ活動という2種類のガンマ活動が発見された。誘発ガンマ活動は、リズムの各音の開始後に確認された。この活動は位相 同期(ピークと谷が音の正確な開始と直接関係している)していることが判明しており、リズムにギャップ(拍子抜け)がある場合には現れない。誘導ガンマ活 動は位相同期が見られなかったが、各拍動と一致していることが分かった。しかし、誘導ガンマ活動はリズムにギャップがある場合でも収まらなかったことか ら、誘導ガンマ活動は聴覚入力とは無関係に、一種の内部メトロノームとして機能している可能性があることが示された。 |
| Tonality Tonality describes the relationships between the elements of melody and harmony – tones, intervals, chords, and scales. These relationships are often characterized as hierarchical, such that one of the elements dominates or attracts another. They occur both within and between every type of element, creating a rich and time-varying perception between tones and their melodic, harmonic, and chromatic contexts. In one conventional sense, tonality refers to just the major and minor scale types – examples of scales whose elements are capable of maintaining a consistent set of functional relationships. The most important functional relationship is that of the tonic note (the first note in a scale) and the tonic chord (the first note in the scale with the third and fifth note) with the rest of the scale. The tonic is the element which tends to assert its dominance and attraction over all others, and it functions as the ultimate point of attraction, rest and resolution for the scale.[18] The right auditory cortex is primarily involved in perceiving pitch, and parts of harmony, melody and rhythm.[16] One study by Petr Janata found that there are tonality-sensitive areas in the medial prefrontal cortex, the cerebellum, the superior temporal sulci of both hemispheres and the superior temporal gyri (which has a skew towards the right hemisphere).[19] Hemispheric asymmetries in the processing of dissonant/consonant sounds have been demonstrated. ERP studies have shown larger evoked responses over the left temporal area in response to dissonant chords, and over the right one, in response to consonant chords.[20] |
調性 調性は、旋律と和音の要素間の関係を説明するものである。音、音程、和音、音階などである。これらの関係は、一方の要素が他方を支配したり引きつけたりす るといった、階層的な関係として特徴づけられることが多い。これらの関係は、あらゆる種類の要素の内部および要素間で発生し、音色と、その旋律、和音、半 音階的な文脈との間に、時間とともに変化する豊かな知覚を生み出す。一般的な意味では、調性とは長音階と短音階を指す。音階の要素が機能的な関係を一貫し て維持できる音階の例である。最も重要な機能的関係は、主音(スケールの最初の音)と主和音(スケールの最初の音と第3音および第5音)と、それ以外のス ケールとの関係である。主音は、他のすべての音に対して支配力と魅力を主張する傾向にある要素であり、スケールにとって究極の引力、休息、解決のポイント として機能する。 右の聴覚野は主に音の高さの知覚に関与しており、和声、旋律、リズムの一部も知覚する。[16] ペトル・ヤナタによるある研究では、内側前頭前皮質、小脳、両半球の上側頭溝、上側頭回(右半球にやや偏っている)に調性感受領域があることが分かった。 [19] 調和する音/調和しない音の処理における半球の非対称性は実証されている。ERP研究では、調和しない和音に対する反応では左側頭領域でより大きな誘発電 位が、調和する和音に対する反応では右側頭領域でより大きな誘発電位が示されている。[20] |
| Music production and performance Motor control functions Musical performance usually involves at least three elementary motor control functions: timing, sequencing, and spatial organization of motor movements. Accuracy in timing of movements is related to musical rhythm. Rhythm, the pattern of temporal intervals within a musical measure or phrase, in turn creates the perception of stronger and weaker beats.[21] Sequencing and spatial organization relate to the expression of individual notes on a musical instrument. These functions and their neural mechanisms have been investigated separately in many studies, but little is known about their combined interaction in producing a complex musical performance.[21] The study of music requires examining them together. |
音楽の制作と演奏 運動制御機能 音楽の演奏には通常、タイミング、シーケンス、運動の空間的構成という少なくとも3つの基本的な運動制御機能が関わっている。運動のタイミングの正確さは 音楽のリズムと関係している。リズムは、音楽の小節やフレーズ内の時間間隔のパターンであり、強拍と弱拍の知覚を生み出す。シーケンスと空間的構成は、楽 器の個々の音符の表現と関係している。 これらの機能と神経メカニズムは、多くの研究で個別に調査されてきたが、複雑な音楽パフォーマンスを生み出す際のそれらの相互作用についてはほとんど知られていない。[21] 音楽の研究では、それらを一緒に調べる必要がある。 |
| Timing Although neural mechanisms involved in timing movement have been studied rigorously over the past 20 years, much remains controversial. The ability to phrase movements in precise time has been accredited to a neural metronome or clock mechanism where time is represented through oscillations or pulses.[22][23][24][25] An opposing view to this metronome mechanism has also been hypothesized stating that it is an emergent property of the kinematics of movement itself.[24][25][26] Kinematics is defined as parameters of movement through space without reference to forces (for example, direction, velocity and acceleration).[21] Functional neuroimaging studies, as well as studies of brain-damaged patients, have linked movement timing to several cortical and sub-cortical regions, including the cerebellum, basal ganglia and supplementary motor area (SMA).[21] Specifically the basal ganglia and possibly the SMA have been implicated in interval timing at longer timescales (1 second and above), while the cerebellum may be more important for controlling motor timing at shorter timescales (milliseconds).[22][27] Furthermore, these results indicate that motor timing is not controlled by a single brain region, but by a network of regions that control specific parameters of movement and that depend on the relevant timescale of the rhythmic sequence.[21] |
タイミング 運動のタイミングに関わる神経メカニズムは過去20年以上にわたって厳密に研究されてきたが、依然として論争の的となっている部分も多い。運動を正確なタ イミングで表現する能力は、振動やパルスによって時間を表現する神経メトロノームまたは時計メカニズムによるものであると認められている。[22] [23][24][25] このメトロノームメカニズムに対する反対意見も仮説として提唱されており、それは運動の運動力学自体の創発的性質であるというものである。 [24][25][26] 運動学とは、力(例えば、方向、速度、加速度)を参照せずに空間における運動のパラメータとして定義される。[21] 機能的ニューロイメージング研究や脳損傷患者の研究により、運動のタイミングは小脳、大脳基底核、補足運動野(SMA)を含むいくつかの皮質および皮質下 の領域に関連していることが分かっている。 [21] 具体的には、大脳基底核と、おそらくは補足運動野(SMA)が、より長い時間軸(1秒以上)における間隔タイミングに関与していると考えられている。一 方、小脳はより短い時間軸(ミリ秒)における運動タイミングの制御に重要である可能性がある。 [22][27] さらに、これらの結果は、運動タイミングは単一の脳領域ではなく、運動の特定のパラメータを制御する領域のネットワークによって制御されており、そのリズ ムシーケンスの関連時間スケールに依存していることを示している。[21] |
| Sequencing Motor sequencing has been explored in terms of either the ordering of individual movements, such as finger sequences for key presses, or the coordination of subcomponents of complex multi-joint movements.[21] Implicated in this process are various cortical and sub-cortical regions, including the basal ganglia, the SMA and the pre-SMA, the cerebellum, and the premotor and prefrontal cortices, all involved in the production and learning of motor sequences but without explicit evidence of their specific contributions or interactions amongst one another.[21] In animals, neurophysiological studies have demonstrated an interaction between the frontal cortex and the basal ganglia during the learning of movement sequences.[28] Human neuroimaging studies have also emphasized the contribution of the basal ganglia for well-learned sequences.[29] The cerebellum is arguably important for sequence learning and for the integration of individual movements into unified sequences,[29][30][31][32][33] while the pre-SMA and SMA have been shown to be involved in organizing or chunking of more complex movement sequences.[34][35] Chunking, defined as the re-organization or re-grouping of movement sequences into smaller sub-sequences during performance, is thought to facilitate the smooth performance of complex movements and to improve motor memory.[21] Lastly, the premotor cortex has been shown to be involved in tasks that require the production of relatively complex sequences, and it may contribute to motor prediction.[36][37] |
シーケンス モーターシーケンスは、キーを押す際の指の動きなどの個々の動きの順序、または複雑な多関節運動のサブコンポーネントの調整という観点から研究されてき た。 [21] このプロセスに関与しているのは、大脳基底核、SMAおよび前SMA、小脳、運動前野および前頭前野皮質など、さまざまな皮質および皮質下の領域である。 これらはすべて運動シーケンスの生成と学習に関与しているが、それらの特定の貢献や相互の相互作用に関する明確な証拠はない。 [21] 動物では、神経生理学的研究により、運動シーケンスの学習中に前頭皮質と大脳基底核との間に相互作用があることが実証されている。[28] ヒトの神経画像研究でも、よく学習されたシーケンスに対する大脳基底核の寄与が強調されている。[29] 小脳は、シーケンス学習と個々の動きを統一されたシーケンスに統合する上で重要であると考えられているが、[29][30][31][32][33] 一方、前SMAとSMAは、より複雑な動きのシーケンスの組織化やチャンキングに関与していることが示されている。 [34][35] チャンキングとは、運動の実行中に、運動シーケンスをより小さなサブシーケンスに再編成または再グループ化することと定義され、複雑な運動の円滑な実行を 促進し、運動記憶を向上させるものと考えられている。[21] 最後に、運動前野は比較的複雑なシーケンスの生成を必要とする課題に関与することが示されており、運動予測にも寄与している可能性がある。[36] [37] |
| Spatial organization Few studies of complex motor control have distinguished between sequential and spatial organization, yet expert musical performances demand not only precise sequencing but also spatial organization of movements. Studies in animals and humans have established the involvement of parietal, sensory–motor and premotor cortices in the control of movements, when the integration of spatial, sensory and motor information is required.[38][39] Few studies so far have explicitly examined the role of spatial processing in the context of musical tasks. |
空間的構成 複雑な運動制御に関する研究の多くは、シーケンシャルな組織と空間的な組織を区別してはこなかったが、熟練した音楽演奏には正確なシーケンスだけでなく、 動きの空間的な組織も求められる。動物と人間を対象とした研究により、空間、感覚、運動の情報の統合が必要な場合、運動制御には頭頂葉、感覚運動野、運動 前野が関与することが明らかになっている。[38][39] 音楽的なタスクにおける空間処理の役割を明確に調査した研究は、これまでほとんどない。 |
| Auditory-motor interactions Feedforward and feedback interactions An auditory–motor interaction may be loosely defined as any engagement of or communication between the two systems. Two classes of auditory-motor interaction are "feedforward" and "feedback".[21] In feedforward interactions, it is the auditory system that predominately influences the motor output, often in a predictive way.[40] An example is the phenomenon of tapping to the beat, where the listener anticipates the rhythmic accents in a piece of music. Another example is the effect of music on movement disorders: rhythmic auditory stimuli have been shown to improve walking ability in Parkinson's disease and stroke patients.[41][42] Feedback interactions are particularly relevant in playing an instrument such as a violin, or in singing, where pitch is variable and must be continuously controlled. If auditory feedback is blocked, musicians can still execute well-rehearsed pieces, but expressive aspects of performance are affected.[43] When auditory feedback is experimentally manipulated by delays or distortions,[44] motor performance is significantly altered: asynchronous feedback disrupts the timing of events, whereas alteration of pitch information disrupts the selection of appropriate actions, but not their timing. This suggests that disruptions occur because both actions and percepts depend on a single underlying mental representation.[21] |
聴覚と運動の相互作用 フィードフォワードとフィードバックの相互作用 聴覚と運動の相互作用とは、大まかに定義すると、この2つのシステム間の関与またはコミュニケーションを指す。聴覚と運動の相互作用には、「フィードフォ ワード」と「フィードバック」の2種類がある。[21] フィードフォワードの相互作用では、主に聴覚システムが運動出力に影響を及ぼす。多くの場合、予測的な方法で影響を及ぼす。[40] 例としては、音楽のリズムのアクセントを聴き手が予測して、それに合わせてタップする現象が挙げられる。別の例としては、運動障害に対する音楽の影響があ る。リズミカルな聴覚刺激は、パーキンソン病や脳卒中の患者の歩行能力を改善することが示されている。[41][42] フィードバックの相互作用は、バイオリンなどの楽器演奏や歌において特に重要である。音の高さは変化しやすく、常に制御する必要がある。聴覚フィードバッ クが遮断された場合、音楽家は十分に練習した曲を演奏することはできるが、パフォーマンスの表現力に影響が出る。[43] 聴覚フィードバックが遅延や歪みによって実験的に操作された場合、[44] 運動パフォーマンスは著しく変化する。非同期フィードバックはイベントのタイミングを崩し、ピッチ情報の変化は適切な行動の選択を妨げるが、タイミングは 妨げない。これは、行動と知覚の両方が単一の基礎となる心的表現に依存しているために混乱が生じることを示唆している。[21] |
| Models of auditory–motor interactions Several models of auditory–motor interactions have been advanced. The model of Hickok and Poeppel,[45] which is specific for speech processing, proposes that a ventral auditory stream maps sounds onto meaning, whereas a dorsal stream maps sounds onto articulatory representations. They and others[46] suggest that posterior auditory regions at the parieto-temporal boundary are crucial parts of the auditory–motor interface, mapping auditory representations onto motor representations of speech, and onto melodies.[47] |
聴覚と運動の相互作用のモデル 聴覚と運動の相互作用のモデルはいくつか提唱されている。 ヒコックとポエペルのモデル[45]は、音声処理に特化しており、腹側聴覚路が音を意味にマッピングする一方で、背側経路が音を構音表現にマッピングする というものである。彼らや他の研究者[46]は、頭頂葉と側頭葉の境界にある後部聴覚領域が、聴覚と運動のインターフェースの重要な部分であり、聴覚表現 を音声の運動表現やメロディにマッピングしていると示唆している。[47] |
| Mirror/echo neurons and auditory–motor interactions The mirror neuron system has an important role in neural models of sensory–motor integration. There is considerable evidence that neurons respond to both actions and the accumulated observation of actions. A system proposed to explain this understanding of actions is that visual representations of actions are mapped onto our own motor system.[48] Some mirror neurons are activated both by the observation of goal-directed actions, and by the associated sounds produced during the action. This suggests that the auditory modality can access the motor system.[49][50] While these auditory–motor interactions have mainly been studied for speech processes, and have focused on Broca's area and the vPMC, as of 2011, experiments have begun to shed light on how these interactions are needed for musical performance. Results point to a broader involvement of the dPMC and other motor areas.[21] The literature has shown a highly specialized cortical network in the skilled musician's brain that codes the relationship between musical gestures and their corresponding sounds. The data hint at the existence of an audiomotor mirror network involving the right superior temporal gyrus, the premotor cortex, the inferior frontal and inferior parietal areas, among other areas.[51] |
ミラーニューロンとエコーニューロン、聴覚と運動の相互作用 ミラーニューロンシステムは、感覚と運動の統合の神経モデルにおいて重要な役割を果たしている。ニューロンが、行動と行動の観察の蓄積の両方に対して反応 するという証拠が数多くある。この行動の理解を説明するシステムとして、行動の視覚的表現が自身の運動システムにマッピングされるというものが提案されて いる。 ミラーニューロンの一部は、目的志向的な行動の観察と、その行動中に発生する関連音の両方によって活性化される。これは、聴覚が運動系にアクセスできるこ とを示唆している。[49][50] これらの聴覚と運動の相互作用は主に言語処理について研究されており、ブローカ野とvPMCに焦点が当てられてきたが、2011年現在、これらの相互作用 が音楽演奏にどのように必要とされるかについて、実験により解明され始めている。その結果、dPMCやその他の運動野のより広範な関与が指摘されている。 [21] 文献では、熟練した音楽家の脳には、音楽的ジェスチャーとそれに対応する音の関係をコード化する高度に専門化された皮質ネットワークが存在することが示さ れている。データからは、右側の上側頭回、運動前野、下前頭野および下頭頂葉などの領域を含む、聴覚運動ミラーネットワークの存在が示唆されている。 [51] |
| Music and language See also: Musical semantics and Musical syntax Certain aspects of language and melody have been shown to be processed in near identical functional brain areas. Brown, Martinez and Parsons (2006) examined the neurological structural similarities between music and language.[52] Utilizing positron emission tomography (PET), the findings showed that both linguistic and melodic phrases produced activation in almost identical functional brain areas. These areas included the primary motor cortex, supplementary motor area, Broca's area, anterior insula, primary and secondary auditory cortices, temporal pole, basal ganglia, ventral thalamus and posterior cerebellum. Differences were found in lateralization tendencies as language tasks favoured the left hemisphere, but the majority of activations were bilateral which produced significant overlap across modalities.[52] Syntactical information mechanisms in both music and language have been shown to be processed similarly in the brain. Jentschke, Koelsch, Sallat and Friederici (2008) conducted a study investigating the processing of music in children with specific language impairments (SLI).[53] Children with typical language development (TLD) showed ERP patterns different from those of children with SLI, which reflected their challenges in processing music-syntactic regularities. Strong correlations between the ERAN (Early Right Anterior Negativity—a specific ERP measure) amplitude and linguistic and musical abilities provide additional evidence for the relationship of syntactical processing in music and language.[53] However, production of melody and production of speech may be subserved by different neural networks. Stewart, Walsh, Frith and Rothwell (2001) studied the differences between speech production and song production using transcranial magnetic stimulation (TMS).[54] Stewart et al. found that TMS applied to the left frontal lobe disturbs speech but not melody supporting the idea that they are subserved by different areas of the brain. The authors suggest that a reason for the difference is that speech generation can be localized well but the underlying mechanisms of melodic production cannot. Alternatively, it was also suggested that speech production may be less robust than melodic production and thus more susceptible to interference.[54] Language processing is a function more of the left side of the brain than the right side, particularly Broca's area and Wernicke's area, though the roles played by the two sides of the brain in processing different aspects of language are still unclear. Music is also processed by both the left and the right sides of the brain.[52][55] Recent evidence further suggest shared processing between language and music at the conceptual level.[56] It has also been found that, among music conservatory students, the prevalence of absolute pitch is much higher for speakers of tone language, even controlling for ethnic background, showing that language influences how musical tones are perceived.[57][58] |
音楽と言語 参照:音楽の意味論と音楽の統語論 言語と旋律の特定の側面は、ほぼ同一の機能的脳領域で処理されることが示されている。Brown、Martinez、Parsons(2006)は、音楽 と言語の神経構造上の類似性を調査した。[52] 陽電子放射断層撮影(PET)を利用した調査の結果、言語的および旋律的なフレーズの両方が、ほぼ同一の機能的脳領域で活性化することが示された。これら の領域には、一次運動野、補足運動野、ブローカ野、前部島皮質、一次および二次聴覚野、側頭極、大脳基底核、腹側視床、後部小脳が含まれる。言語課題では 左半球が優位となる傾向に違いが見られたが、大半の活性化は両側性であり、様式を越えて著しい重複が見られた。 音楽と言語における構文情報のメカニズムは、脳内で同様に処理されることが示されている。Jentschke、Koelsch、Sallat、 Friederici(2008年)は、特定言語障害(SLI)を持つ子供における音楽の処理を調査する研究を実施した。[53] 典型的な言語発達(TLD)を持つ子供は、SLIを持つ子供とは異なるERPパターンを示し、これは音楽の構文規則性を処理する際に困難を抱えていること を反映している。ERAN(早期右前部陰性電位:特定のERP測定値)の振幅と言語能力および音楽能力との間に強い相関関係があることは、音楽と言語にお ける統語処理の関係を示すさらなる証拠となる。[53] しかし、メロディの生成と音声の生成は、異なる神経ネットワークによって行われている可能性がある。スチュワート、ウォルシュ、フリス、ロスウェル (2001年)は、経頭蓋磁気刺激(TMS)を用いて、音声生成と歌の生成の違いを研究した。[54] スチュワートらは、左前頭葉にTMSを適用すると音声は妨害されるが、メロディは妨害されないことを発見し、音声とメロディが脳の異なる領域によって処理 されているという考えを裏付けた。著者らは、音声生成は局在化できるが、メロディ生成の根本的なメカニズムは局在化できないことが、この違いの原因である と示唆している。あるいは、音声生成は旋律生成よりも頑強性が低く、干渉を受けやすいという可能性も示唆されている。 言語処理は、右脳よりも左脳、特にブローカ野とウェルニッケ野の機能である。しかし、言語の異なる側面を処理する際の脳の両半球の役割については、まだ不 明な点が多い。音楽もまた、脳の左半球と右半球の両方で処理される。[52][55] 最近の証拠はさらに、言語と音楽の概念レベルでの処理の共通性を示唆している。[56] また、音楽院の学生の間では、母語がトーン言語である場合、民族背景を考慮しても、絶対音感を持つ人の割合がはるかに高いことが分かっており、言語が音楽 の音色がどのように知覚されるかに影響を及ぼしていることが示されている。[57][58] |
Musician vs. non-musician processing Professional pianists show less cortical activation for complex finger movement tasks due to structural differences in the brain. Differences Brain structure within musicians and non-musicians is distinctly different. Gaser and Schlaug (2003) compared brain structures of professional musicians with non-musicians and discovered gray matter volume differences in motor, auditory and visual-spatial brain regions.[59] Specifically, positive correlations were discovered between musician status (professional, amateur and non-musician) and gray matter volume in the primary motor and somatosensory areas, premotor areas, anterior superior parietal areas and in the inferior temporal gyrus bilaterally. This strong association between musician status and gray matter differences supports the notion that musicians' brains show use-dependent structural changes.[60] Due to the distinct differences in several brain regions, it is unlikely that these differences are innate but rather due to the long-term acquisition and repetitive rehearsal of musical skills. Brains of musicians also show functional differences from those of non-musicians. Krings, Topper, Foltys, Erberich, Sparing, Willmes and Thron (2000) utilized fMRI to study brain area involvement of professional pianists and a control group while performing complex finger movements.[61] Krings et al. found that the professional piano players showed lower levels of cortical activation in motor areas of the brain. It was concluded that a lesser amount of neurons needed to be activated for the piano players due to long-term motor practice which results in the different cortical activation patterns. Koeneke, Lutz, Wustenberg and Jancke (2004) reported similar findings in keyboard players.[62] Skilled keyboard players and a control group performed complex tasks involving unimanual and bimanual finger movements. During task conditions, strong hemodynamic responses in the cerebellum were shown by both non-musicians and keyboard players, but non-musicians showed the stronger response. This finding indicates that different cortical activation patterns emerge from long-term motor practice. This evidence supports previous data showing that musicians require fewer neurons to perform the same movements. Musicians have been shown to have significantly more developed left planum temporales, and have also shown to have a greater word memory.[63] Chan's study controlled for age, grade point average and years of education and found that when given a 16 word memory test, the musicians averaged one to two more words above their non musical counterparts. |
音楽家と非音楽家の処理能力 プロのピアニストは、脳の構造上の違いにより、複雑な指の動きの課題に対する大脳皮質の活性化が少ない。 違い 音楽家と非音楽家では、脳の構造が明らかに異なる。GaserとSchlaug(2003年)は、プロの音楽家と音楽家でない人の脳構造を比較し、運動、 聴覚、視覚空間に関連する脳領域に灰白質容積の違いがあることを発見した。[59] 具体的には、音楽家(プロ、アマチュア、音楽家でない人)と一次運動野および体性感覚野、運動前野、上側頭回前方、両側の下側頭回における灰白質容積との 間に正の相関関係があることが判明した。音楽家であることと灰白質の違いとの間にこのような強い関連性があることは、音楽家の脳は使用依存性の構造変化を 示すという考え方を裏付けるものである。[60] いくつかの脳領域に明確な違いがあることから、これらの違いは生得的なものではなく、むしろ音楽的スキルの長期的な習得と反復練習によるものである可能性 が高い。 また、音楽家の脳は、音楽家でない人の脳とは機能的に異なることも分かっている。Krings、Topper、Foltys、Erberich、 Sparing、Willmes、Thron(2000年)は、fMRI(機能的磁気共鳴画像法)を用いて、複雑な指の動きを行っているときのプロのピア ニストと対照群の脳領域の関与について研究した。Kringsらは、プロのピアニストは脳の運動領域における皮質の活性化レベルが低いことを発見した。長 期間の運動練習により、ピアノ奏者にはより少ない数のニューロンを活性化させるだけでよいことが結論づけられ、その結果、異なる皮質活性化パターンが生じ ると考えられる。Koeneke、Lutz、Wustenberg、Jancke(2004年)は、鍵盤奏者についても同様の結果を報告している。 [62]熟練した鍵盤奏者と対照群が、単手および両手の指の動きを伴う複雑なタスクを行った。課題遂行中、非音楽家とキーボード奏者の両者で小脳における 強い血流動態反応が示されたが、非音楽家の方がより強い反応を示した。この結果は、長期間の運動練習により異なる皮質活性化パターンが生じることを示して いる。この証拠は、音楽家が同じ動きを行うのに必要なニューロン数が少ないことを示す過去のデータを裏付けるものである。 音楽家は左側頭葉平面が著しく発達しており、また単語記憶力も高いことが示されている。[63] チャンの研究では、年齢、成績平均値、教育年数を統制した上で、16語の単語記憶テストを行ったところ、音楽家は非音楽家よりも平均して1~2語多く記憶 していたことが分かった。 |
| Similarities Studies have shown that the human brain has an implicit musical ability.[64][65] Koelsch, Gunter, Friederici and Schoger (2000) investigated the influence of preceding musical context, task relevance of unexpected chords and the degree of probability of violation on music processing in both musicians and non-musicians.[64] Findings showed that the human brain unintentionally extrapolates expectations about impending auditory input. Even in non-musicians, the extrapolated expectations are consistent with music theory. The ability to process information musically supports the idea of an implicit musical ability in the human brain. In a follow-up study, Koelsch, Schroger, and Gunter (2002) investigated whether ERAN and N5 could be evoked preattentively in non-musicians.[65] Findings showed that both ERAN and N5 can be elicited even in a situation where the musical stimulus is ignored by the listener indicating that there is a highly differentiated preattentive musicality in the human brain. |
類似点 研究により、人間の脳には潜在的な音楽能力があることが示されている。[64][65] Koelsch、Gunter、Friederici、Schoger(2000年)は、音楽家と音楽家でない人の両方について、先行する音楽的コンテク スト、予期せぬ和音のタスク関連性、音楽処理における違反の確率の度合いが及ぼす影響を調査した。[64] その結果、人間の脳は、差し迫った聴覚入力について、意図せずに予測を外挿することが示された。音楽家でない人でも、その推測された期待は音楽理論と一致 している。音楽的に情報を処理する能力は、人間の脳に潜在的な音楽能力があるという考え方を裏付けるものである。その後の研究で、Koelsch、 Schroger、Gunter(2002年)は、音楽家ではない被験者において、ERANとN5が注意を要する前に誘発されるかどうかを調査した。 [65] その結果、音楽的な刺激が聞き手によって無視されている状況でも、ERANとN5の両方が誘発されることが判明した。これは、人間の脳には高度に分化した 注意を要しない音楽性があることを示している。 |
| Gender differences Minor neurological differences regarding hemispheric processing exist between brains of males and females. Koelsch, Maess, Grossmann and Friederici (2003) investigated music processing through EEG and ERPs and discovered gender differences.[66] Findings showed that females process music information bilaterally and males process music with a right-hemispheric predominance. However, the early negativity of males was also present over the left hemisphere. This indicates that males do not exclusively utilize the right hemisphere for musical information processing. In a follow-up study, Koelsch, Grossman, Gunter, Hahne, Schroger and Friederici (2003) found that boys show lateralization of the early anterior negativity in the left hemisphere but found a bilateral effect in girls.[67] This indicates a developmental effect as early negativity is lateralized in the right hemisphere in men and in the left hemisphere in boys. |
性差 男性と女性の脳には、半球処理に関する軽微な神経学的差異が存在する。Koelsch、Maess、Grossmann、Friederici (2003)は、EEGとERPによる音楽処理を調査し、性差を発見した。[66] 調査結果によると、女性は音楽情報を左右両半球で処理し、男性は右半球優位で音楽を処理することが分かった。しかし、男性の初期の陰性電位は左半球にも存 在していた。このことは、男性が音楽情報の処理に右半球だけを使用しているわけではないことを示している。追跡調査では、Koelsch、 Grossman、Gunter、Hahne、Schroger、Friederici(2003年)が、男児では左半球に早期前部負電位の側方化が認め られるが、女児では両側性に認められることを発見した。[67] これは、早期前部負電位が男性では右半球に、男児では左半球に側方化されるという発達的効果を示している。 |
| Handedness differences It has been found that subjects who are lefthanded, particularly those who are also ambidextrous, perform better than righthanders on short term memory for the pitch.[68][69] It was hypothesized that this handedness advantage is due to the fact that lefthanders have more duplication of storage in the two hemispheres than do righthanders. Other work has shown that there are pronounced differences between righthanders and lefthanders (on a statistical basis) in how musical patterns are perceived, when sounds come from different regions of space. This has been found, for example, in the Octave illusion[70][71] and the Scale illusion.[72][73] |
利き手による違い 左利き、特に両利きの人の方が、右利きの人よりも音高の短期記憶において優れたパフォーマンスを発揮することが分かっている。[68][69] この利き手による利点の要因として、左利きの人の方が右利きの人よりも2つの半球における記憶の重複が多いという事実が挙げられるという仮説が立てられて いる。他の研究では、異なる空間領域から音が聞こえる場合、音楽パターンがどのように知覚されるかについて、右利きと左利きには顕著な違いがあることが示 されている(統計的根拠に基づく)。これは、例えばオクターブ錯視[70][71]や音階錯視[72][73]で確認されている。 |
| Musical imagery See also: Audiation Musical imagery refers to the experience of replaying music by imagining it inside the head.[74] Musicians show a superior ability for musical imagery due to intense musical training.[75] Herholz, Lappe, Knief and Pantev (2008) investigated the differences in neural processing of a musical imagery task in musicians and non-musicians. Utilizing magnetoencephalography (MEG), Herholz et al. examined differences in the processing of a musical imagery task with familiar melodies in musicians and non-musicians. Specifically, the study examined whether the mismatch negativity (MMN) can be based solely on imagery of sounds. The task involved participants listening to the beginning of a melody, continuation of the melody in his/her head and finally hearing a correct/incorrect tone as further continuation of the melody. The imagery of these melodies was strong enough to obtain an early preattentive brain response to unanticipated violations of the imagined melodies in the musicians. These results indicate similar neural correlates are relied upon for trained musicians imagery and perception. Additionally, the findings suggest that modification of the imagery mismatch negativity (iMMN) through intense musical training results in achievement of a superior ability for imagery and preattentive processing of music. Perceptual musical processes and musical imagery may share a neural substrate in the brain. A PET study conducted by Zatorre, Halpern, Perry, Meyer and Evans (1996) investigated cerebral blood flow (CBF) changes related to auditory imagery and perceptual tasks.[76] These tasks examined the involvement of particular anatomical regions as well as functional commonalities between perceptual processes and imagery. Similar patterns of CBF changes provided evidence supporting the notion that imagery processes share a substantial neural substrate with related perceptual processes. Bilateral neural activity in the secondary auditory cortex was associated with both perceiving and imagining songs. This implies that within the secondary auditory cortex, processes underlie the phenomenological impression of imagined sounds. The supplementary motor area (SMA) was active in both imagery and perceptual tasks suggesting covert vocalization as an element of musical imagery. CBF increases in the inferior frontal polar cortex and right thalamus suggest that these regions may be related to retrieval and/or generation of auditory information from memory. |
音楽的イメージ 関連情報: 聴覚化 音楽的イメージとは、頭の中で音楽を想像することで音楽を再生する体験を指す。[74] 音楽家は、厳しい音楽トレーニングにより、音楽的イメージに優れた能力を発揮する。[75] Herholz、Lappe、Knief、Pantev(2008年)は、音楽家と音楽家でない人の音楽的イメージタスクにおける神経処理の違いを調査し た。ヘルホルツらは、脳磁図(MEG)を用いて、音楽家と非音楽家における、聞き覚えのあるメロディの音楽的イメージタスクの処理における違いを調査し た。具体的には、この研究では、ミスマッチ陰性電位(MMN)が音のイメージのみに基づいて生じるかどうかを調べた。このタスクでは、参加者はメロディの 冒頭を聞き、頭の中でメロディを続け、最後に正しい音または間違った音をメロディのさらなる続きとして聞く。これらのメロディのイメージは十分に強力で あったため、ミュージシャンが想像したメロディの予期せぬ逸脱に対する早期の前注意的脳反応が得られた。これらの結果は、訓練を受けたミュージシャンのイ メージと知覚には、同様の神経相関が関与していることを示している。さらに、この研究結果は、集中的な音楽トレーニングによるイメージミスマッチ陰性電位 (iMMN)の変化が、優れたイメージ能力と音楽の前注意的処理能力の獲得につながることを示唆している。 音楽の知覚プロセスと音楽的イメージは、脳内の神経基質を共有している可能性がある。Zatorre、Halpern、Perry、Meyer、 Evans(1996)によるPET研究では、聴覚的イメージと知覚的タスクに関連する脳血流(CBF)の変化が調査された。[76] これらのタスクでは、知覚プロセスとイメージの機能的共通性だけでなく、特定の解剖学的領域の関与についても調査された。同様のCBF変化パターンは、イ メージ処理が関連する知覚処理とかなりの神経基質を共有するという考えを裏付ける証拠となった。 二次聴覚野における両側の神経活動は、楽曲の知覚と想像の両方に関連していた。 これは、二次聴覚野において、想像上の音の現象学的印象の根底にあるプロセスを示唆している。 補足運動野(SMA)は、イメージと知覚の両方のタスクで活性化しており、音楽的イメージの要素として潜在的な発声を示唆している。下前頭極皮質および右 視床におけるCBFの増加は、これらの領域が記憶からの聴覚情報の検索および/または生成に関連している可能性を示唆している。 |
| Emotion Main article: Music and emotion Music is able to create an intensely pleasurable experience that can be described as "chills".[77] Blood and Zatorre (2001) used PET to measure changes in cerebral blood flow while participants listened to music that they knew to give them the "chills" or any sort of intensely pleasant emotional response. They found that as these chills increase, many changes in cerebral blood flow are seen in brain regions such as the amygdala, orbitofrontal cortex, ventral striatum, midbrain, and the ventral medial prefrontal cortex. Many of these areas appear to be linked to reward, motivation, emotion, and arousal, and are also activated in other pleasurable situations.[77] The resulting pleasure responses enable the release dopamine, serotonin, and oxytocin. Nucleus accumbens (a part of striatum) is involved in both music related emotions, as well as rhythmic timing. [78] According to the National Institute of Health, children and adults who are suffering from emotional trauma have been able to benefit from the use of music in a variety of ways. The use of music has been essential in helping children who struggle with focus, anxiety, and cognitive function by using music in therapeutic way. Music therapy has also helped children cope with autism, pediatric cancer, and pain from treatments. Emotions induced by music activate similar frontal brain regions compared to emotions elicited by other stimuli.[60] Schmidt and Trainor (2001) discovered that valence (i.e. positive vs. negative) of musical segments was distinguished by patterns of frontal EEG activity.[79] Joyful and happy musical segments were associated with increases in left frontal EEG activity whereas fearful and sad musical segments were associated with increases in right frontal EEG activity. Additionally, the intensity of emotions was differentiated by the pattern of overall frontal EEG activity. Overall frontal region activity increased as affective musical stimuli became more intense.[79] When unpleasant melodies are played, the posterior cingulate cortex activates, which indicates a sense of conflict or emotional pain.[16] The right hemisphere has also been found to be correlated with emotion, which can also activate areas in the cingulate in times of emotional pain, specifically social rejection (Eisenberger). This evidence, along with observations, has led many musical theorists, philosophers and neuroscientists to link emotion with tonality. This seems almost obvious because the tones in music seem like a characterization of the tones in human speech, which indicate emotional content. The vowels in the phonemes of a song are elongated for a dramatic effect, and it seems as though musical tones are simply exaggerations of the normal verbal tonality. |
感情 詳細は「音楽と感情」を参照 音楽は「戦慄」と表現されるような、非常に強い快感を生み出すことができる。[77] BloodとZatorre(2001年)は、参加者が「戦慄」や、あらゆる種類の非常に強い快感を生み出す音楽を聴いている間の脳血流の変化をPETで 測定した。その結果、ゾクゾクするような感覚が高まると、扁桃体、眼窩前頭皮質、腹側被殻、中脳、腹内側前頭前皮質などの脳領域で、脳血流に多くの変化が 現れることが分かった。これらの領域の多くは、報酬、動機付け、感情、覚醒と関連しているように見え、他の快楽的な状況でも活性化される。[77] その結果、快楽反応が起こり、ドーパミン、セロトニン、オキシトシンの放出が促される。側坐核(線状体の一部)は、音楽に関連する感情とリズムのタイミン グの両方に関与している。 [78] 米国保健社会福祉省によると、情緒的外傷に苦しむ子供や大人は、音楽のさまざまな活用法から恩恵を受けている。 集中力や不安、認知機能に問題を抱える子供たちを治療する上で、音楽の活用は不可欠である。 音楽療法は、自閉症、小児がん、治療に伴う痛みに苦しむ子供たちにも役立っている。 音楽によって引き起こされる感情は、他の刺激によって引き起こされる感情と同様に、前頭葉の脳領域を活性化させる。[60] シュミットとトレイナー(2001年)は、音楽のセグメントの価値(すなわち、ポジティブ対ネガティブ)が、前頭葉の脳波活動のパターンによって区別され ることを発見した。[79] 喜びや楽しさを感じさせる音楽のセグメントは、左の前頭葉の脳波活動の増加と関連しており、一方で、恐怖や悲しみを感じさせる音楽のセグメントは、右の前 頭葉の脳波活動の増加と関連していた。さらに、感情の強さは、前頭部全体の脳波活動のパターンによって区別された。感情的な音楽刺激が強くなるにつれ、前 頭部全体の活動が増加した。[79] 不快なメロディが演奏されると、後部帯状皮質が活性化し、葛藤や感情的な苦痛が示される。[16] 右半球も感情と相関関係にあることが判明しており、感情的な苦痛、特に社会的拒絶を経験した際には、帯状皮質の一部が活性化することがある。(アイゼン バーガー) こうした証拠や観察結果から、多くの音楽理論家、哲学者、神経科学者は、感情と調性を結びつけるようになった。音楽の音色が人間の話し言葉の音色の特徴を 表しているように思えるため、これはほぼ明白である。曲の音素の母音はドラマチックな効果を狙って長く伸ばされるが、音楽の音色は通常の話し言葉の音色を 単純に誇張したものにすぎないように思える。 |
| Memory Main article: Music-related memory Neuropsychology of musical memory Musical memory involves both explicit and implicit memory systems.[80] Explicit musical memory is further differentiated between episodic (where, when and what of the musical experience) and semantic (memory for music knowledge including facts and emotional concepts). Implicit memory centers on the 'how' of music and involves automatic processes such as procedural memory and motor skill learning – in other words skills critical for playing an instrument. Samson and Baird (2009) found that the ability of musicians with Alzheimer's Disease to play an instrument (implicit procedural memory) may be preserved. |
記憶 詳細は「音楽に関連する記憶」を参照 音楽の記憶に関する神経心理学 音楽の記憶には、明示的記憶と暗示的記憶の両方が関与している。[80] 明示的な音楽の記憶は、さらにエピソード(音楽体験の場所、時間、内容)と意味(事実や感情的な概念を含む音楽の知識の記憶)に区別される。暗示的記憶は 音楽の「方法」に焦点を当て、手続き記憶や運動技能学習などの自動的なプロセスに関与している。つまり、楽器演奏に不可欠な技能である。Samsonと Baird(2009年)は、アルツハイマー病を患う音楽家の楽器演奏能力(暗黙的な手続き記憶)は維持されている可能性があることを発見した。 |
| Neural correlates of musical memory A PET study looking into the neural correlates of musical semantic and episodic memory found distinct activation patterns.[81] Semantic musical memory involves the sense of familiarity of songs. The semantic memory for music condition resulted in bilateral activation in the medial and orbital frontal cortex, as well as activation in the left angular gyrus and the left anterior region of the middle temporal gyri. These patterns support the functional asymmetry favouring the left hemisphere for semantic memory. Left anterior temporal and inferior frontal regions that were activated in the musical semantic memory task produced activation peaks specifically during the presentation of musical material, suggestion that these regions are somewhat functionally specialized for musical semantic representations. Episodic memory of musical information involves the ability to recall the former context associated with a musical excerpt.[81] In the condition invoking episodic memory for music, activations were found bilaterally in the middle and superior frontal gyri and precuneus, with activation predominant in the right hemisphere. Other studies have found the precuneus to become activated in successful episodic recall.[82] As it was activated in the familiar memory condition of episodic memory, this activation may be explained by the successful recall of the melody. When it comes to memory for pitch, there appears to be a dynamic and distributed brain network subserves pitch memory processes. Gaab, Gaser, Zaehle, Jancke and Schlaug (2003) examined the functional anatomy of pitch memory using functional magnetic resonance imaging (fMRI).[83] An analysis of performance scores in a pitch memory task resulted in a significant correlation between good task performance and the supramarginal gyrus (SMG) as well as the dorsolateral cerebellum. Findings indicate that the dorsolateral cerebellum may act as a pitch discrimination processor and the SMG may act as a short-term pitch information storage site. The left hemisphere was found to be more prominent in the pitch memory task than the right hemispheric regions. |
音楽の記憶における神経相関 音楽の意味記憶とエピソード記憶における神経相関を調査したPET研究では、異なる活性化パターンが発見された。[81] 音楽の意味記憶には、楽曲に対する親近感が関与している。音楽条件における意味記憶では、内側および眼窩前頭皮質の左右両側での活性化、および左角回と側 頭葉回中央部の左前方領域での活性化が見られた。これらのパターンは、意味記憶において左半球が優位となる機能的非対称性を裏付けるものである。音楽の意 味記憶タスクで活性化された左側頭前野および下前頭前野は、音楽素材の提示中に特に活性化のピークを生み出した。このことは、これらの領域が音楽の意味表 現に対してある程度機能的に特化されていることを示唆している。 音楽情報のエピソード記憶には、音楽の一部に関連する以前の文脈を思い出す能力が関与している。[81] 音楽のエピソード記憶を呼び起こす条件では、左右の中前頭回および上前頭回、ならびに楔前部で活性化が認められ、活性化は右半球で優位であった。他の研究 では、成功したエピソード想起において、Precuneusが活性化することが分かっている。[82] エピソード記憶の馴染みのある記憶条件で活性化していることから、この活性化はメロディの想起が成功したことによるものと考えられる。 音高の記憶に関しては、音高の記憶プロセスを司る、ダイナミックで分散的な脳ネットワークが存在するようだ。Gaab、Gaser、Zaehle、 Jancke、Schlaug(2003)は、機能的磁気共鳴画像法(fMRI)を用いて音高記憶の機能解剖学を調査した。[83] 音高記憶課題におけるパフォーマンススコアの分析により、課題のパフォーマンスが良好な場合、上側頭回(SMG)および小脳の外側後頭部との間に有意な相 関があることが判明した。この結果から、小脳の背外側部は音高弁別処理装置として、また上側頭回は音高の短期記憶の保存場所として機能している可能性があ ることが示唆される。音高記憶課題では、右半球の領域よりも左半球の領域がより顕著であることが分かった。 |
| Therapeutic effects of music on memory Musical training has been shown to aid memory. Altenmuller et al. studied the difference between active and passive musical instruction and found both that over a longer (but not short) period of time, the actively taught students retained much more information than the passively taught students. The actively taught students were also found to have greater cerebral cortex activation. The passively taught students weren't wasting their time; they, along with the active group, displayed greater left hemisphere activity, which is typical in trained musicians.[84] Research suggests we listen to the same songs repeatedly because of musical nostalgia. One major study, published in the journal Memory & Cognition, found that music enables the mind to evoke memories of the past, known as music-evoked autobiographical memories.[85] |
音楽が記憶に与える治療効果 音楽の訓練は記憶力を向上させることが分かっている。Altenmullerらは、能動的音楽指導と受動的音楽指導の違いを研究し、どちらも長期間(短時 間ではない)にわたって、能動的に指導された学生の方が受動的に指導された学生よりも多くの情報を記憶していることを発見した。能動的に指導された学生は 大脳皮質の活性化もより高いことが分かった。受動的に教えられた生徒たちは時間を無駄にしていたわけではなく、能動的に教えられたグループと同様に左半球 の活動が活発であり、これは訓練を受けた音楽家によく見られる現象である。 研究によると、私たちは音楽的な郷愁から同じ曲を繰り返し聴くという。学術誌『Memory & Cognition』に掲載されたある主要な研究では、音楽は過去の記憶を呼び起こすことを可能にし、音楽によって引き起こされる自伝的記憶として知られ ていることが分かった。 |
| Attention Treder et al.[86] identified neural correlates of attention when listening to simplified polyphonic music patterns. In a musical oddball experiment, they had participants shift selective attention to one out of three different instruments in music audio clips, with each instrument occasionally playing one or several notes deviating from an otherwise repetitive pattern. Contrasting attended versus unattended instruments, ERP analysis shows subject- and instrument-specific responses including P300 and early auditory components. The attended instrument could be classified offline with high accuracy. This indicates that attention paid to a particular instrument in polyphonic music can be inferred from ongoing EEG, a finding that is potentially relevant for building more ergonomic music-listing based brain-computer interfaces.[86] |
注意 Treder ら[86]は、単純化された多声音楽パターンを聴く際の注意の神経相関を特定した。音楽のオドボール実験では、参加者は音楽オーディオクリップの3つの異 なる楽器のうち1つに選択的に注意を移し、各楽器は時折、それ以外の反復パターンから逸脱した1つまたは複数の音を演奏した。注意を向けた楽器と向けな かった楽器を比較したERP分析では、P300や初期の聴覚成分を含む、被験者および楽器特有の反応が示された。注意が向けられた楽器は、オフラインで高 い精度で分類できる。これは、多声音楽における特定の楽器に対する注意が、進行中の脳波から推測できることを示しており、より人間工学的な音楽リスニング に基づくブレイン・コンピュータ・インターフェースの構築に役立つ可能性がある。[86] |
| Development Musical four-year-olds have been found to have one greater left hemisphere intrahemispheric coherence.[84] Musicians have been found to have more developed anterior portions of the corpus callosum in a study by Cowell et al. in 1992. This was confirmed by a study by Schlaug et al. in 1995 that found that classical musicians between the ages of 21 and 36 have significantly greater anterior corpora callosa than the non-musical control. Schlaug also found that there was a strong correlation of musical exposure before the age of seven, and a great increase in the size of the corpus callosum.[84] These fibers join together the left and right hemispheres and indicate an increased relaying between both sides of the brain. This suggests the merging between the spatial- emotiono-tonal processing of the right brain and the linguistical processing of the left brain. This large relaying across many different areas of the brain might contribute to music's ability to aid in memory function. |
発達 音楽的才能を持つ4歳児は、左半球の内部における結合性が1つ多いことが分かっている。[84] 1992年のカウェルらによる研究では、音楽家は脳梁の前部がより発達していることが分かっている。これは、1995年のSchlaugらの研究によって も確認されており、21歳から36歳までのクラシック音楽家は、音楽家ではない対照群と比較して、前交連が有意に大きいことが分かった。また、 Schlaugは7歳以前に音楽に触れていたことと、脳梁のサイズの大幅な増加との間に強い相関関係があることも発見した。[84] これらの繊維は左右の大脳半球をつないでおり、脳の両側間の連絡が増加していることを示している。これは右脳の空間的・情動的・調性的処理と左脳の言語的 処理が融合していることを示唆している。脳の多くの異なる領域にわたるこの大きな中継が、音楽が記憶機能を助ける能力に寄与しているのかもしれない。 |
| Impairment Main article: Music-specific disorders Focal hand dystonia Focal hand dystonia is a task-related movement disorder associated with occupational activities that require repetitive hand movements.[87] Focal hand dystonia is associated with abnormal processing in the premotor and primary sensorimotor cortices. An fMRI study examined five guitarists with focal hand dystonia.[88] The study reproduced task-specific hand dystonia by having guitarists use a real guitar neck inside the scanner as well as performing a guitar exercise to trigger abnormal hand movement. The dystonic guitarists showed significantly more activation of the contralateral primary sensorimotor cortex as well as a bilateral underactivation of premotor areas. This activation pattern represents abnormal recruitment of the cortical areas involved in motor control. Even in professional musicians, widespread bilateral cortical region involvement is necessary to produce complex hand movements such as scales and arpeggios. The abnormal shift from premotor to primary sensorimotor activation directly correlates with guitar-induced hand dystonia. |
障害 詳細は「音楽特有の障害」を参照 局所性手指ジストニア 局所性手ジストニアは、反復的な手の動きを必要とする職業活動に関連する、作業関連運動障害である。[87] 局所性手ジストニアは、運動前野および一次感覚運動野における異常な処理に関連している。fMRI研究では、局所性手ジストニアを持つ5人のギタリストを 調査した。[88] この研究では、ギタリストにスキャナー内で本物のギターネックを使わせ、ギターの練習を行わせることで、作業特異的な手ジストニアを再現した。ジストニア を患うギタリストは、反対側の一次感覚運動野の著しい活性化と、両側の運動前野の低活性化を示した。この活性化パターンは、運動制御に関与する皮質領域の 異常な動員を表している。プロのミュージシャンであっても、音階やアルペジオなどの複雑な手の動きを生み出すには、広範囲にわたる両側の皮質領域の関与が 必要である。運動前野から一次感覚運動野への異常な活性化シフトは、ギター演奏による手のジストニアと直接相関している。 |
| Music agnosia Music agnosia, an auditory agnosia, is a syndrome of selective impairment in music recognition.[89] Three cases of music agnosia are examined by Dalla Bella and Peretz (1999); C.N., G.L., and I.R.. All three of these patients suffered bilateral damage to the auditory cortex which resulted in musical difficulties while speech understanding remained intact. Their impairment is specific to the recognition of once familiar melodies. They are spared in recognizing environmental sounds and in recognizing lyrics. Peretz (1996) has studied C.N.'s music agnosia further and reports an initial impairment of pitch processing and spared temporal processing.[90] C.N. later recovered in pitch processing abilities but remained impaired in tune recognition and familiarity judgments. Musical agnosias may be categorized based on the process which is impaired in the individual.[91] Apperceptive music agnosia involves an impairment at the level of perceptual analysis involving an inability to encode musical information correctly. Associative music agnosia reflects an impaired representational system which disrupts music recognition. Many of the cases of music agnosia have resulted from surgery involving the middle cerebral artery. Patient studies have surmounted a large amount of evidence demonstrating that the left side of the brain is more suitable for holding long-term memory representations of music and that the right side is important for controlling access to these representations. Associative music agnosias tend to be produced by damage to the left hemisphere, while apperceptive music agnosia reflects damage to the right hemisphere. |
音楽失認 音楽失認、聴覚失認は、音楽認識における選択的障害の症候群である。[89] ダラ・ベラとペレツ(1999年)は、音楽失認の3つの症例を調査した。C.N.、G.L.、I.R.の3人の患者は、いずれも聴覚野に両側性の損傷を 負っており、その結果、音楽の理解に困難が生じているが、音声の理解には問題がない。彼らの障害は、かつて耳にしたことのあるメロディを認識することに特 異的に現れる。環境音の認識や歌詞の認識には障害は見られない。Peretz(1996)は、C.N.の音楽失認についてさらに研究を進め、初期にはピッ チ処理に障害が見られ、時間処理には障害が見られないことを報告している。[90] C.N.は後にピッチ処理能力は回復したが、曲の認識と馴染みのある音楽の判断には障害が残った。 音楽性認知障害は、障害の生じているプロセスに基づいて分類することができる。[91] 知覚性音楽性認知障害は、音楽情報の正確なエンコードができない知覚分析レベルでの障害を伴う。連想性音楽性認知障害は、音楽の認識を妨げる表象システム の障害を反映している。音楽失認の症例の多くは、中大脳動脈に関わる手術が原因である。患者を対象とした研究では、音楽の長期記憶の表象を保持するには左 脳が適しており、これらの表象へのアクセスを制御するには右脳が重要であることを示す多くの証拠が示されている。連想音楽失認は左半球の損傷によって生じ ることが多く、知覚音楽失認は右半球の損傷によって生じる。 |
| Congenital amusia Congenital amusia, otherwise known as tone deafness, is a term for lifelong musical problems which are not attributable to intellectual disability, lack of exposure to music or deafness, or brain damage after birth.[92] Amusic brains have been found in fMRI studies to have less white matter and thicker cortex than controls in the right inferior frontal cortex. These differences suggest abnormal neuronal development in the auditory cortex and inferior frontal gyrus, two areas which are important in musical-pitch processing. Studies on those with amusia suggest different processes are involved in speech tonality and musical tonality. Congenital amusics lack the ability to distinguish between pitches and so are for example unmoved by dissonance and playing the wrong key on a piano. They also cannot be taught to remember a melody or to recite a song; however, they are still capable of hearing the intonation of speech, for example, distinguishing between "You speak French" and "You speak French?" when spoken. |
先天性音痴 先天性音痴、別名音痴は、知的障害、音楽への接触不足、難聴、または出生後の脳障害に起因しない生涯にわたる音楽的問題を指す用語である。[92] fMRI研究では、音痴の脳では、右の下前頭皮質において、対照群よりも白質が少なく、皮質が厚いことが分かっている。これらの違いは、音楽のピッチ処理 において重要な役割を果たす聴覚野と下前頭回において、神経細胞の異常な発達が起こっていることを示唆している。 音楽音痴の研究では、音声の音調と音楽の音調には異なるプロセスが関与していることが示唆されている。先天性音楽音痴の人は音の高さを区別する能力に欠け ているため、例えば、不協和音に感動しないし、ピアノで違うキーを弾いても動じない。また、メロディを覚えたり歌を暗誦したりすることも教えられない。し かし、彼らは話し言葉のイントネーションを聞き取る能力はあり、例えば「あなたはフランス語を話しますね」と「あなたはフランス語を話しますか」という言 葉を区別することができる。 |
Amygdala damage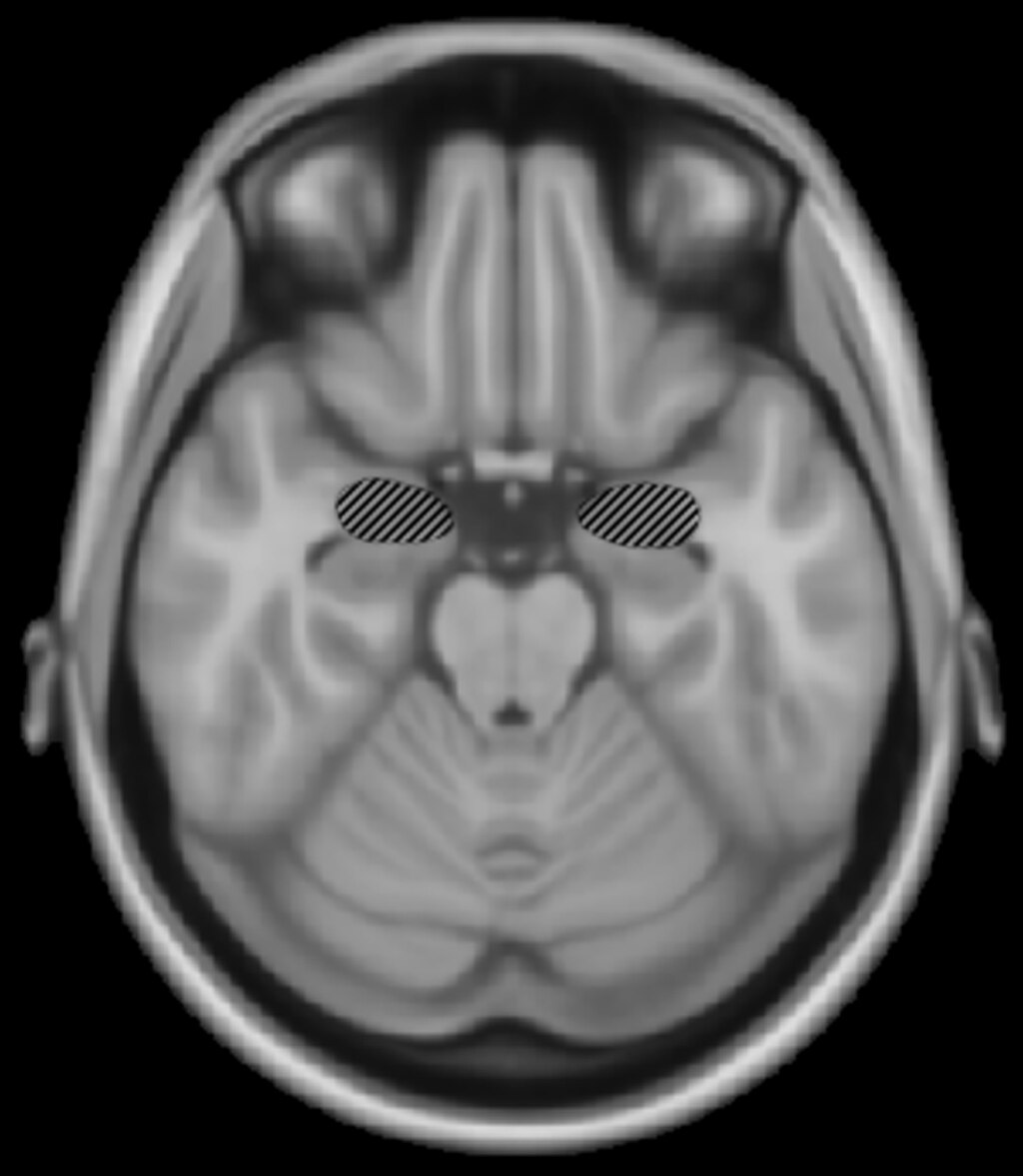 Damage to the amygdala may impair recognition of scary music. Damage to the amygdala has selective emotional impairments on musical recognition. Gosselin, Peretz, Johnsen and Adolphs (2007) studied S.M., a patient with bilateral damage of the amygdala with the rest of the temporal lobe undamaged and found that S.M. was impaired in recognition of scary and sad music.[93] S.M.'s perception of happy music was normal, as was her ability to use cues such as tempo to distinguish between happy and sad music. It appears that damage specific to the amygdala can selectively impair recognition of scary music. |
扁桃体の損傷 扁桃体の損傷は、怖い音楽の認識を損なう可能性がある。 扁桃体の損傷は、音楽の認識における感情の選択的障害を引き起こす。Gosselin、Peretz、Johnsen、Adolphs(2007年)は、 側頭葉の他の部分は損傷していないものの、扁桃体の両側が損傷している患者S.M.を研究し、S.M.は怖い音楽や悲しい音楽の認識に障害があることを発 見した。[93] S.M.の楽しい音楽の知覚は正常であり、テンポなどの手がかりを使って楽しい音楽と悲しい音楽を区別する能力も正常であった。扁桃体特有の損傷は、怖い 音楽の認識を特異的に損なう可能性がある。 |
| Selective deficit in music reading Specific musical impairments may result from brain damage leaving other musical abilities intact. Cappelletti, Waley-Cohen, Butterworth and Kopelman (2000) studied a single case study of patient P.K.C., a professional musician who sustained damage to the left posterior temporal lobe as well as a small right occipitotemporal lesion.[94] After sustaining damage to these regions, P.K.C. was selectively impaired in the areas of reading, writing and understanding musical notation but maintained other musical skills. The ability to read aloud letters, words, numbers and symbols (including musical ones) was retained. However, P.K.C. was unable to read aloud musical notes on the staff regardless of whether the task involved naming with the conventional letter or by singing or playing. Yet despite this specific deficit, P.K.C. retained the ability to remember and play familiar and new melodies. |
音楽の読譜における選択的欠損 特定の音楽的障害は、他の音楽的能力には影響を与えない脳障害によって生じる可能性がある。Cappelletti、Waley-Cohen、 Butterworth、Kopleman(2000年)は、左側後部側頭葉および右側後頭側頭葉に小さな病変を持つプロの音楽家である患者P.K.C. の単一ケーススタディを行った。[94] これらの領域に損傷を受けた後、P.K.C.は楽譜の読み書きと理解の分野で選択的に障害を受けたが、その他の音楽能力は維持していた。文字、単語、数 字、記号(音楽記号を含む)を音読する能力は維持されていた。しかし、P.K.C.は、従来の文字による名称、歌、演奏のいずれの課題においても、五線譜 上の音符を音読することができなかった。しかし、この特定の欠陥にもかかわらず、P.K.C.は、馴染みのあるメロディや新しいメロディを記憶し演奏する 能力は維持していた。 |
| Auditory arrhythmia Arrhythmia in the auditory modality is defined as a disturbance of rhythmic sense; and includes deficits such as the inability to rhythmically perform music, the inability to keep time to music and the inability to discriminate between or reproduce rhythmic patterns.[95] A study investigating the elements of rhythmic function examined Patient H.J., who acquired arrhythmia after sustaining a right temporoparietal infarct.[95] Damage to this region impaired H.J.'s central timing system which is essentially the basis of his global rhythmic impairment. H.J. was unable to generate steady pulses in a tapping task. These findings suggest that keeping a musical beat relies on functioning in the right temporal auditory cortex. |
聴覚性不整脈 聴覚における不整脈は、リズム感覚の障害と定義され、音楽のリズムを正確に演奏できない、音楽に合わせて拍子を刻めない、リズムパターンを識別したり再現 したりできないなどの欠損が含まれる。 [95] リズム機能の要素を調査した研究では、右側頭頭頂葉梗塞を発症した後、不整脈を患った患者H.J.を調査した。[95] この領域の損傷により、H.J.の中心タイミングシステムが損なわれた。このシステムは、本質的にはH.J.の全体的なリズム障害の基礎となるものであ る。H.J.はタッピング課題において安定したパルスを生成することができなかった。これらの知見は、音楽のビートを維持するには右側頭聴覚野の機能が重 要であることを示唆している。 |
| https://en.wikipedia.org/wiki/Neuroscience_of_music |
リ ンク
文 献
そ の他の情報
Copyleft,
CC, Mitzub'ixi Quq Chi'j, 1996-2099
Copyleft, CC, Mitzub'ixi Quq Chi'j, 1996-2099
![]()
☆
 ☆
☆