脳中心主義
Brain-centrism
![]()
☆ 「脳中心主義」または「大脳中心主義(cerebrocentrism)」とは、意識、主観、人間の経験の主な源泉であり、その場であるのは脳であるという考え方を指し、しばしば脳が思考、感情、行動の唯一の決定要因であることを暗示する。
| "Brain-centrism"
or "cerebrocentrism" refers to the idea that the brain is the primary
source and locus of consciousness, subjectivity, and human experience,
often implying that the brain is the sole determinant of our thoughts,
feelings, and behaviors. Here's a more detailed explanation: Core Idea: Brain-centrism posits that the brain is the ultimate seat of our being, with all aspects of our experience originating within it. Alternative Perspectives: Some argue that this view is too narrow and that our experiences are also shaped by the body, environment, and social interactions, emphasizing the importance of embodiment and circular interaction. Examples of Brain-Centric Concepts: Brain-centric Design: This approach focuses on how we acquire knowledge and uses neuroscience to present information for deep understanding. Brain-Centric Reliability System (BCR): This system focuses on organizational safety and reliability, emphasizing the importance of human performance and decision-making. Brain-centric Model of Suicidal Behavior: This model focuses on internal stressors and the role of mental health conditions in suicidal behavior. Criticisms of Brain-Centrism: Embodiment: The idea that our experiences are solely determined by the brain ignores the body's role in shaping our perceptions and behaviors. Circular Interaction: Consciousness and experience are not simply the product of brain processes but also result from the continuous interaction between the brain, body, and environment. Living Processes: The brain is not a computer, but an organ of a living person that is involved in circular, living processes. |
「脳中心主義」または「大脳中心主義」とは、意識、主観、人間の経験の主な源泉であり、その場であるのは脳であるという考え方を指し、しばしば脳が思考、感情、行動の唯一の決定要因であることを暗示する。 以下に、より詳細な説明を示す。 中心となる考え方: 脳中心主義は、脳が人間の存在の究極の座であり、人間の経験のあらゆる側面は脳内で生じるという考えである。 代替的な視点: この見方はあまりにも狭すぎるとの意見もあり、人間の経験は身体、環境、社会的相互作用によっても形作られると主張する人もいる。身体化と循環的相互作用の重要性を強調する。 脳中心主義の概念の例: 脳中心主義のデザイン: このアプローチは、私たちが知識を習得する方法に焦点を当て、深い理解を得るために神経科学を利用して情報を提示する。 脳中心の信頼性システム(BCR):このシステムは、組織の安全性と信頼性に焦点を当て、人間のパフォーマンスと意思決定の重要性を強調する。 脳中心の自殺行動モデル:このモデルは、自殺行動における内的なストレス要因と精神保健状態の役割に焦点を当てる。 脳中心主義への批判: 身体化:人間の経験は脳によってのみ決定されるという考え方は、人間の知覚や行動の形成における身体の役割を無視している。 循環的相互作用:意識や経験は、脳のプロセスによる単純な産物ではなく、脳、身体、環境の間の継続的な相互作用の結果でもある。 生命プロセス:脳はコンピュータではなく、循環的な生命プロセスに関与する生きた人格の器官である。 |
 京都府立医科大学の老年神経科学研究室の英語ホームページ. |
 Principal fissures and lobes of the cerebrum viewed laterally. Figure 728 from Gray's Anatomy with labels removed. |
| The
cerebrum (pl.: cerebra), telencephalon or endbrain[1] is the largest
part of the brain, containing the cerebral cortex (of the two cerebral
hemispheres) as well as several subcortical structures, including the
hippocampus, basal ganglia, and olfactory bulb. In the human brain, the
cerebrum is the uppermost region of the central nervous system. The
cerebrum develops prenatally from the forebrain (prosencephalon). In
mammals, the dorsal telencephalon, or pallium, develops into the
cerebral cortex, and the ventral telencephalon, or subpallium, becomes
the basal ganglia. The cerebrum is also divided into approximately
symmetric left and right cerebral hemispheres. With the assistance of the cerebellum, the cerebrum controls all voluntary actions in the human body. |
大
脳(複数形:大脳)または終脳[1]は、脳の最も大きな部分であり、大脳皮質(2つの大脳半球)のほか、海馬、大脳基底核、嗅球などの皮質下構造を含む。
ヒトの脳において、大脳は中枢神経系の最上位の領域である。大脳は胎児期に前脳(前脳)から発達する。哺乳類では、背側終脳(大脳皮質)が大脳皮質に、腹
側終脳(大脳皮質下)が大脳基底核に発達する。大脳はまた、左右ほぼ対称の左大脳半球と右大脳半球に分かれている。 小脳の補助を受け、大脳は人体のすべての随意運動を制御している。 |
Structure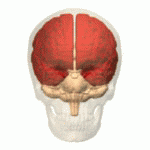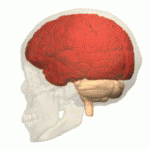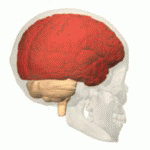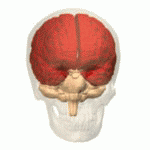 Location of the human cerebrum (red). The cerebrum is the largest part of the brain. Depending upon the position of the animal, it lies either in front or on top of the brainstem. In humans, the cerebrum is the largest and best-developed of the five major divisions of the brain. The cerebrum is made up of the two cerebral hemispheres and their cerebral cortices (the outer layers of grey matter), and the underlying regions of white matter.[2] Its subcortical structures include the hippocampus, basal ganglia and olfactory bulb. The cerebrum consists of two C-shaped cerebral hemispheres, separated from each other by a deep fissure called the longitudinal fissure. |
構造 ヒトの大脳の位置(赤)。 大脳は脳の中で最も大きな部分である。動物の位置によって、大脳は脳幹の前または上に位置する。ヒトでは、大脳は脳の5つの主要な部分の中で最も大きく、最も発達している。 大脳は2つの大脳半球とその大脳皮質(灰白質の表層部)、およびその下の白質から構成されている。[2] 皮質下の構造には、海馬、大脳基底核、嗅球がある。大脳は2つのC字形の大脳半球からなり、縦裂と呼ばれる深い裂によって互いに隔てられている。 |
Cerebral cortex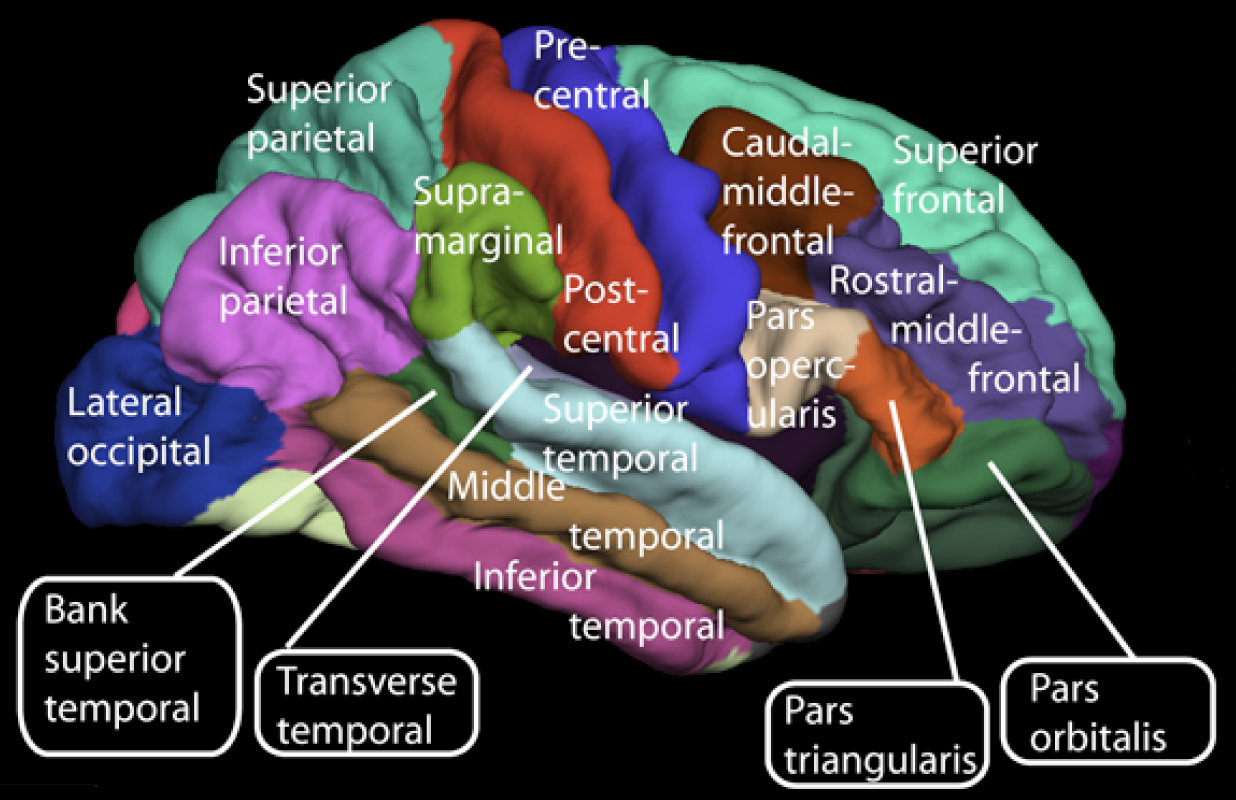 Surface of the cerebrum The cerebral cortex, the outer layer of grey matter of the cerebrum, is found only in mammals. In larger mammals, including humans, the surface of the cerebral cortex folds to create gyri (ridges) and sulci (furrows) which increase the surface area.[3] The cerebral cortex is generally classified into four lobes: the frontal, parietal, occipital and temporal lobes. The lobes are classified based on their overlying neurocranial bones.[4] A smaller lobe is the insular lobe, a part of the cerebral cortex folded deep within the lateral sulcus that separates the temporal lobe from the parietal and frontal lobes, is located within each hemisphere of the mammalian brain. |
大脳皮質 大脳の表面 大脳皮質は、大脳の灰白質の外層であり、哺乳類にのみ存在する。ヒトを含む大型哺乳類では、大脳皮質の表面が折り重なって溝(脳回)と溝(脳溝)が形成され、表面積が増加する。 大脳皮質は一般的に4つの葉に分類される。すなわち、前頭葉、頭頂葉、後頭葉、側頭葉である。これらの葉は、覆いかぶさる神経頭蓋骨に基づいて分類され る。4つより小さい葉は島葉であり、側頭葉と頭頂葉および前頭葉を隔てる外側溝の奥深くに折り重なる大脳皮質の一部分である。島葉は、哺乳類の脳の各半球 内に位置している。 |
| Cerebral hemispheres The cerebrum is divided by the medial longitudinal fissure into two cerebral hemispheres, the right and the left. The cerebrum is contralaterally organized, i.e., the right hemisphere controls and processes signals (predominantly) from the left side of the body, while the left hemisphere controls and processes signals (predominantly) from the right side of the body.[4] According to current knowledge, this is due to an axial twist that occurs in the early embryo.[5] There is a strong but not complete bilateral symmetry between the hemispheres, while lateralization tends to increase with increasing brain size.[6] The lateralization of brain function looks at the known and possible differences between the two. |
大脳半球 大脳は内側縦裂によって右と左の2つの大脳半球に分けられる。大脳は反対側と対応するように構成されており、すなわち、右半球は主に身体の左側からの信号 を制御し処理し、左半球は主に身体の右側からの信号を制御し処理する。 [4] 現在の知見によると、これは初期の胚に軸捻転が起こるためである。[5] 大脳半球の間には完全ではないが強い左右対称性がある。また、脳のサイズが大きくなるにつれて、機能の局在化は増加する傾向にある。[6] 脳機能の局在化は、2つの間の既知および可能性のある相違を調査するものである。 |
| Development In the developing vertebrate embryo, the neural tube is subdivided into four unseparated sections which then develop further into distinct regions of the central nervous system; these are the prosencephalon (forebrain), the mesencephalon (midbrain) the rhombencephalon (hindbrain) and the spinal cord.[7] The prosencephalon develops further into the telencephalon and the diencephalon. The dorsal telencephalon gives rise to the pallium (cerebral cortex in mammals and reptiles) and the ventral telencephalon generates the basal ganglia. The diencephalon develops into the thalamus and hypothalamus, including the optic vesicles (future retina).[8] The dorsal telencephalon then forms two lateral telencephalic vesicles, separated by the midline, which develop into the left and right cerebral hemispheres. Birds and fish have a dorsal telencephalon, like all vertebrates, but it is generally unlayered and therefore not considered a cerebral cortex. Only a layered cytoarchitecture can be considered a cortex. |
発生 脊椎動物の胚が発達する過程において、神経管は4つの未分離のセクションに細分化され、その後、中枢神経系の異なる領域へとさらに発達する。これらは前脳 (prosencephalon)、中脳(mesencephalon)、後脳(rhombencephalon)、脊髄である。前脳はさらに終脳 (telencephalon)と間脳(diencephalon)へと発達する。背側終脳は大脳皮質(哺乳類および爬虫類)を生じ、腹側終脳は線条体を 生じる。間脳は視蓋(将来の網膜)を含む視床と視床下部へと発達する。[8] 背側終脳はその後、正中線によって隔てられた2つの側方終脳小脳小体(lateral telencephalic vesicles)を形成し、これらは左右の大脳半球へと発達する。鳥類や魚類は、他の脊椎動物と同様に、背側終脳を持つが、一般的に層を持たないため、 大脳皮質とは見なされない。層を持つ細胞構築のみが、皮質と見なされる。 |
| Functions Note: As the cerebrum is a gross division with many subdivisions and sub-regions, it is important to state that this section lists only functions that the cerebrum as a whole serves. (See main articles on cerebral cortex and basal ganglia for more information.) The cerebrum is a major part of the brain, controlling emotions, hearing, vision, personality and much more. It controls all precision of voluntary actions, and it functions as the center of sensory perception, memory, thoughts and judgement; the cerebrum also functions as the center of voluntary motor activities. Motor functions Upper motor neurons in the primary motor cortex send their axons to the brainstem and spinal cord to synapse on the lower motor neurons, which innervate the muscles. Damage to motor areas of the cortex can lead to certain types of motor neuron disease. This kind of damage results in loss of muscular power and precision rather than total paralysis. Sensory processing The primary sensory areas of the cerebral cortex receive and process visual, auditory, somatosensory, gustatory, and olfactory information. Together with association cortical areas, these brain regions synthesize sensory information into our perceptions of the world. Olfaction Main article: Sense of smell The olfactory bulb, responsible for the sense of smell, takes up a large area of the cerebrum in most vertebrates. However, in humans, this part of the brain is much smaller and lies underneath the frontal lobe. The olfactory sensory system is unique since the neurons in the olfactory bulb send their axons directly to the olfactory cortex, rather than to the thalamus first. Olfaction is also the only sense that is represented by the ipsilateral side of the brain. Damage to the olfactory bulb results in a loss of olfaction (the sense of smell). Language and communication Main article: Language Speech and language are mainly attributed to parts of the cerebral cortex. Motor portions of language are attributed to Broca's area within the frontal lobe. Speech comprehension is attributed to Wernicke's area, at the temporal-parietal lobe junction. These two regions are interconnected by a large white matter tract, the arcuate fasciculus. Damage to the Broca's area results in expressive aphasia (non-fluent aphasia) while damage to Wernicke's area results in receptive aphasia (also called fluent aphasia). Learning and memory Main article: Memory Explicit or declarative (factual) memory formation is attributed to the hippocampus and associated regions of the medial temporal lobe. This association was originally described after a patient known as HM had both his left and right hippocampus surgically removed to treat chronic [temporal lobe epilepsy]. After surgery, HM had anterograde amnesia, or the inability to form new memories. Implicit or procedural memory, such as complex motor behaviors, involves the basal ganglia. Short-term or working memory involves association areas of the cortex, especially the dorsolateral prefrontal cortex, as well as the hippocampus. |
機能 注:大脳は多くの細分化された部分と亜領域を持つ大まかな区分であるため、このセクションでは大脳全体が果たす機能のみを記載していることを明記しておく ことが重要である。(詳細は大脳皮質と大脳基底核の主記事を参照のこと)大脳は脳の主要な部分であり、感情、聴覚、視覚、人格など、さまざまなものを制御 している。随意運動のあらゆる精密さを制御し、感覚知覚、記憶、思考、判断の中心として機能する。大脳は随意運動の中心としても機能する。 運動機能 一次運動野の上位運動ニューロンは、脳幹と脊髄に軸索を送り、下位運動ニューロンとシナプスを形成し、筋肉に神経を供給する。大脳皮質の運動野が損傷する と、特定の運動ニューロン疾患を引き起こす可能性がある。このような損傷は、完全な麻痺というよりも、筋力の低下や正確性の喪失をもたらす。 感覚処理 大脳皮質の一次感覚野は、視覚、聴覚、体性感覚、味覚、嗅覚の情報を受け取り処理する。連合皮質領域とともに、これらの脳領域は感覚情報を統合し、世界に対する知覚を作り出す。 嗅覚 詳細は「嗅覚」を参照 嗅覚をつかさどる嗅球は、ほとんどの脊椎動物において大脳の大部分を占めている。しかし、ヒトではこの部分ははるかに小さく、前頭葉の下にある。嗅覚の神 経系は独特であり、嗅球のニューロンは視床を経由せずに直接嗅覚皮質に軸索を伸ばしている。嗅覚はまた、脳の同側で表現される唯一の感覚でもある。嗅球に 損傷が生じると、嗅覚(嗅覚)が失われる。 言語とコミュニケーション 詳細は「言語」を参照 言語と会話は主に大脳皮質の部分に起因する。言語の運動部分は前頭葉内のブローカ領域に起因する。言語の理解は側頭葉と頭頂葉の接合部にあるウェルニッケ 領域に起因する。この2つの領域は、大脳白質の束である弓状束によって相互に接続されている。ブローカ領域が損傷すると表出性失語症(非流暢性失語症)と なり、ウェルニッケ領域が損傷すると受容性失語症(流暢性失語症とも呼ばれる)となる。 学習と記憶 詳細は「記憶」を参照 明示的または宣言的(事実)記憶の形成は、海馬および側頭葉内側に関連する領域に起因する。この関連性は、HMとして知られる患者が慢性の側頭葉てんかん を治療するために、左右の海馬を外科的に除去された後に初めて説明された。手術後、HMは順行性健忘症、すなわち新しい記憶を形成できない状態となった。 複雑な運動行動などの暗黙的または手続き的記憶は、大脳基底核が関与する。 短期記憶または作業記憶には、海馬のほか、大脳皮質の連合野、特に背外側前頭前野が関与している。 |
| Other animals In the most primitive vertebrates, the hagfishes and lampreys, the cerebrum is a relatively simple structure receiving nerve impulses from the olfactory bulb. In cartilaginous and lobe-finned fishes and also in amphibians, a more complex structure is present, with the cerebrum being divided into three distinct regions. The lowermost (or ventral) region forms the basal nuclei, and contains fibres connecting the rest of the cerebrum to the thalamus. Above this, and forming the lateral part of the cerebrum, is the paleopallium, while the uppermost (or dorsal) part is referred to as the archipallium. The cerebrum remains largely devoted to olfactory sensation in these animals, in contrast to its much wider range of functions in amniotes.[9] In ray-finned fishes, the structure is somewhat different. The inner surfaces of the lateral and ventral regions of the cerebrum bulge up into the ventricles; these include both the basal nuclei and the various parts of the pallium and may be complex in structure, especially in teleosts. The dorsal surface of the cerebrum is membranous, and does not contain any nervous tissue.[9] In the amniotes, the cerebrum becomes increasingly large and complex. In reptiles, the paleopallium is much larger than in amphibians and its growth has pushed the basal nuclei into the central regions of the cerebrum. As in the lower vertebrates, the grey matter is generally located beneath the white matter, but in some reptiles, it spreads out to the surface to form a primitive cortex, especially in the anterior part of the brain.[9] In mammals, this development proceeds further, so that the cortex covers almost the whole of the cerebral hemispheres, especially in more developed species, such as the primates. The paleopallium is pushed to the ventral surface of the brain, where it becomes the olfactory lobes, while the archipallium becomes rolled over at the medial dorsal edge to form the hippocampus. In placental mammals, a corpus callosum also develops, further connecting the two hemispheres. The complex convolutions of the cerebral surface (see gyrus, gyrification) are also found only in higher mammals.[9] Although some large mammals (such as elephants) have particularly large cerebra, dolphins are the only species (other than humans) to have cerebra accounting for as much as 2 percent of their body weight.[10] The cerebra of birds are similarly enlarged to those of mammals, by comparison with reptiles. The increased size of bird brains was classically attributed to enlarged basal ganglia, with the other areas remaining primitive, but this view has been largely abandoned.[11] Birds appear to have undergone an alternate process of encephalization,[12] as they diverged from the other archosaurs, with few clear parallels to that experienced by mammals and their therapsid ancestors. |
その他の動物 最も原始的な脊椎動物であるメクラウナギやヤツメウナギでは、大脳は比較的単純な構造で、嗅球から神経インパルスを受け取る。軟骨魚類や有葉魚類、両生類 では、より複雑な構造が存在し、大脳は3つの異なる領域に分かれている。最下層(腹側)の領域は基底核を形成し、大脳の残りの部分と視床を結ぶ繊維を含ん でいる。この上方に位置し、大脳の外側部分を形成しているのは古皮質であり、最も上方(または背側)の部分は大脳皮質と呼ばれる。大脳は、有羊膜類では機 能の範囲がはるかに広いのに対し、これらの動物では嗅覚にほぼ特化している。[9] 条鰭類では、その構造はいくらか異なる。大脳の側面および腹面の内部表面は膨らんで脳室を形成しており、そこには基底核とさまざまな皮質部分の両方が含まれている。特に硬骨魚類では、その構造は複雑である可能性がある。大脳の背面は膜状であり、神経組織は含まれていない。 有羊膜類では、大脳はますます大きくなり、複雑になる。爬虫類では、古皮質は両生類よりもはるかに大きく、その成長により基底核は大脳の中央領域に押しや られている。下等脊椎動物と同様に、灰白質は一般的に白質の下に位置しているが、一部の爬虫類では、灰白質は表面に広がり、原始的な皮質を形成している。 特に脳の前部でその傾向が強い。 哺乳類では、この発達はさらに進み、大脳皮質が大脳半球のほぼ全体を覆うようになる。特に霊長類などの発達した種ではその傾向が強い。古皮質は脳の腹側表 面に押しやられ、嗅覚葉となる。一方、原皮質は背側内側で折れ曲がり、海馬を形成する。胎盤を持つ哺乳類では、脳梁も発達し、2つの半球をさらに結びつけ る。大脳皮質の複雑な溝(回、回状化を参照)も、高等哺乳類にのみ見られる。[9] 象などの大型哺乳類の中には特に大脳が大きいものもいるが、体重の2パーセントにも及ぶ大脳を持つのはイルカ(人間以外では)のみである。[10] 鳥類の大脳も、爬虫類と比較すると哺乳類と同様に拡大している。鳥類の脳の拡大は、従来は基底核の拡大によるものとされ、他の領域は原始的なままであると 考えられていたが、この見方はほぼ放棄されている。[11] 鳥類は、他のアーコサウルス類から分岐した際、哺乳類とその祖先である獣弓類が経験したこととはほとんど類似点のない、別の脳化のプロセスを経たようであ る。[12] |
| List of regions in the human brain |
|
| https://en.wikipedia.org/wiki/Cerebrum |
リ ンク
文 献
そ の他の情報
Copyleft, CC, Mitzub'ixi Quq Chi'j, 1996-2099
![]()
☆
 ☆
☆