
生態学
Ecology
☆ 生態学(せいたいがく、英: Ecology)[A]とは、人間を含む生物とその物理的環境との関係に関する自然科学である。生態学は、個体、個体群、群集、生態系、生物圏のレベルで 生物を考察する。生態学は、密接に関連する生物地理学、進化生物学、遺伝学、倫理学、博物学と重複している。 生態学は生物学の一分野であり、環境との関連における生物の存在量、バイオマス、分布を研究する学問である。生命プロセス、相互作用、適応、生物群集を通 じた物質とエネルギーの移動、生態系の後継的発展、種内および種間の協力、競争、捕食、生物多様性のパターンと生態系プロセスへの影響などが含まれる。 生態学は、保全生物学、湿地管理、天然資源管理(農業生態学、農業、林業、農林業、漁業、鉱業、観光業)、都市計画(都市生態学)、地域社会の健康、経済 学、基礎科学および応用科学、人間の社会的相互作用(人間生態学)において実際に応用されている。 エコロジー(ドイツ語:Ökologie)という言葉は、1866年にドイツの科学者エルンスト・ヘッケルによって作られた。今日のような生態学は、 1890年代にアメリカの植物学者のグループによって始まった[1]。適応と自然淘汰に関する進化論的概念は、現代の生態学理論の基礎となっている。 生態系は、生物、それらが構成する群集、およびそれらの環境の非生物的(abiotic)構成要素からなる動的相互作用システムである。一次生産、栄養循 環、ニッチ構築などの生態系プロセスは、環境中のエネルギーと物質の流束を調節する。生態系には生物物理学的フィードバック機構があり、地球の生物(生物 学的)構成要素と非生物(生物学的)構成要素に作用するプロセスを調整する。生態系は生命維持機能を維持し、バイオマス生産(食料、燃料、繊維、医薬 品)、気候の調節、地球規模の生物地球化学的循環、水のろ過、土壌形成、浸食防止、洪水防止、その他科学的、歴史的、経済的、あるいは本質的価値のある多 くの自然特性などの生態系サービスを提供している。
☆エコロジーリンク
++
| Ecology (from
Ancient Greek οἶκος (oîkos) 'house', and -λογία (-logía) 'study of')[A]
is the natural science of the relationships among living organisms,
including humans, and their physical environment. Ecology considers
organisms at the individual, population, community, ecosystem, and
biosphere levels. Ecology overlaps with the closely related sciences of
biogeography, evolutionary biology, genetics, ethology, and natural
history. Ecology is a branch of biology, and is the study of abundance, biomass, and distribution of organisms in the context of the environment. It encompasses life processes, interactions, and adaptations; movement of materials and energy through living communities; successional development of ecosystems; cooperation, competition, and predation within and between species; and patterns of biodiversity and its effect on ecosystem processes. Ecology has practical applications in conservation biology, wetland management, natural resource management (agroecology, agriculture, forestry, agroforestry, fisheries, mining, tourism), urban planning (urban ecology), community health, economics, basic and applied science, and human social interaction (human ecology). The word ecology (German: Ökologie) was coined in 1866 by the German scientist Ernst Haeckel. The science of ecology as we know it today began with a group of American botanists in the 1890s.[1] Evolutionary concepts relating to adaptation and natural selection are cornerstones of modern ecological theory. Ecosystems are dynamically interacting systems of organisms, the communities they make up, and the non-living (abiotic) components of their environment. Ecosystem processes, such as primary production, nutrient cycling, and niche construction, regulate the flux of energy and matter through an environment. Ecosystems have biophysical feedback mechanisms that moderate processes acting on living (biotic) and abiotic components of the planet. Ecosystems sustain life-supporting functions and provide ecosystem services like biomass production (food, fuel, fiber, and medicine), the regulation of climate, global biogeochemical cycles, water filtration, soil formation, erosion control, flood protection, and many other natural features of scientific, historical, economic, or intrinsic value. |
生態学(せいたいがく、英:
Ecology)[A]とは、人間を含む生物とその物理的環境との関係に関する自然科学である。生態学は、個体、個体群、群集、生態系、生物圏のレベルで
生物を考察する。生態学は、密接に関連する生物地理学、進化生物学、遺伝学、倫理学、博物学と重複している。 生態学は生物学の一分野であり、環境との関連における生物の存在量、バイオマス、分布を研究する学問である。生命プロセス、相互作用、適応、生物群集を通 じた物質とエネルギーの移動、生態系の後継的発展、種内および種間の協力、競争、捕食、生物多様性のパターンと生態系プロセスへの影響などが含まれる。 生態学は、保全生物学、湿地管理、天然資源管理(農業生態学、農業、林業、農林業、漁業、鉱業、観光業)、都市計画(都市生態学)、地域社会の健康、経済 学、基礎科学および応用科学、人間の社会的相互作用(人間生態学)において実際に応用されている。 エコロジー(ドイツ語:Ökologie)という言葉は、1866年にドイツの科学者エルンスト・ヘッケルによって作られた。今日のような生態学は、 1890年代にアメリカの植物学者のグループによって始まった[1]。適応と自然淘汰に関する進化論的概念は、現代の生態学理論の基礎となっている。 生態系は、生物、それらが構成する群集、およびそれらの環境の非生物的(abiotic)構成要素からなる動的相互作用システムである。一次生産、栄養循 環、ニッチ構築などの生態系プロセスは、環境中のエネルギーと物質の流束を調節する。生態系には生物物理学的フィードバック機構があり、地球の生物(生物 学的)構成要素と非生物(生物学的)構成要素に作用するプロセスを調整する。生態系は生命維持機能を維持し、バイオマス生産(食料、燃料、繊維、医薬 品)、気候の調節、地球規模の生物地球化学的循環、水のろ過、土壌形成、浸食防止、洪水防止、その他科学的、歴史的、経済的、あるいは本質的価値のある多 くの自然特性などの生態系サービスを提供している。 |
| Levels, scope, and scale of organization Main article: Outline of ecology The scope of ecology contains a wide array of interacting levels of organization spanning micro-level (e.g., cells) to a planetary scale (e.g., biosphere) phenomena. Ecosystems, for example, contain abiotic resources and interacting life forms (i.e., individual organisms that aggregate into populations which aggregate into distinct ecological communities). Because ecosystems are dynamic and do not necessarily follow a linear successional route, changes might occur quickly or slowly over thousands of years before specific forest successional stages are brought about by biological processes. An ecosystem's area can vary greatly, from tiny to vast. A single tree is of little consequence to the classification of a forest ecosystem, but is critically relevant to organisms living in and on it.[2] Several generations of an aphid population can exist over the lifespan of a single leaf. Each of those aphids, in turn, supports diverse bacterial communities.[3] The nature of connections in ecological communities cannot be explained by knowing the details of each species in isolation, because the emergent pattern is neither revealed nor predicted until the ecosystem is studied as an integrated whole.[4] Some ecological principles, however, do exhibit collective properties where the sum of the components explain the properties of the whole, such as birth rates of a population being equal to the sum of individual births over a designated time frame.[5] The main subdisciplines of ecology, population (or community) ecology and ecosystem ecology, exhibit a difference not only in scale but also in two contrasting paradigms in the field. The former focuses on organisms' distribution and abundance, while the latter focuses on materials and energy fluxes.[6] Hierarchy See also: Biological organisation and Biological classification System behaviors must first be arrayed into different levels of the organization. Behaviors corresponding to higher levels occur at slow rates. Conversely, lower organizational levels exhibit rapid rates. For example, individual tree leaves respond rapidly to momentary changes in light intensity, CO2 concentration, and the like. The growth of the tree responds more slowly and integrates these short-term changes. O'Neill et al. (1986)[7]: 76 The scale of ecological dynamics can operate like a closed system, such as aphids migrating on a single tree, while at the same time remaining open about broader scale influences, such as atmosphere or climate. Hence, ecologists classify ecosystems hierarchically by analyzing data collected from finer scale units, such as vegetation associations, climate, and soil types, and integrate this information to identify emergent patterns of uniform organization and processes that operate on local to regional, landscape, and chronological scales. To structure the study of ecology into a conceptually manageable framework, the biological world is organized into a nested hierarchy, ranging in scale from genes, to cells, to tissues, to organs, to organisms, to species, to populations, to guilds, to communities, to ecosystems, to biomes, and up to the level of the biosphere.[8] This framework forms a panarchy[9] and exhibits non-linear behaviors; this means that "effect and cause are disproportionate, so that small changes to critical variables, such as the number of nitrogen fixers, can lead to disproportionate, perhaps irreversible, changes in the system properties."[10]: 14 Biodiversity Main article: Biodiversity Biodiversity refers to the variety of life and its processes. It includes the variety of living organisms, the genetic differences among them, the communities and ecosystems in which they occur, and the ecological and evolutionary processes that keep them functioning, yet ever-changing and adapting. Noss & Carpenter (1994)[11]: 5 Biodiversity (an abbreviation of "biological diversity") describes the diversity of life from genes to ecosystems and spans every level of biological organization. The term has several interpretations, and there are many ways to index, measure, characterize, and represent its complex organization.[12][13][14] Biodiversity includes species diversity, ecosystem diversity, and genetic diversity and scientists are interested in the way that this diversity affects the complex ecological processes operating at and among these respective levels.[13][15][16] Biodiversity plays an important role in ecosystem services which by definition maintain and improve human quality of life.[14][17][18] Conservation priorities and management techniques require different approaches and considerations to address the full ecological scope of biodiversity. Natural capital that supports populations is critical for maintaining ecosystem services[19][20] and species migration (e.g., riverine fish runs and avian insect control) has been implicated as one mechanism by which those service losses are experienced.[21] An understanding of biodiversity has practical applications for species and ecosystem-level conservation planners as they make management recommendations to consulting firms, governments, and industry.[22] Habitat Main article: Habitat Biodiversity of a coral reef. Corals adapt to and modify their environment by forming calcium carbonate skeletons. This provides growing conditions for future generations and forms a habitat for many other species.[23] Long-tailed broadbill building its nest The habitat of a species describes the environment over which a species is known to occur and the type of community that is formed as a result.[24] More specifically, "habitats can be defined as regions in environmental space that are composed of multiple dimensions, each representing a biotic or abiotic environmental variable; that is, any component or characteristic of the environment related directly (e.g. forage biomass and quality) or indirectly (e.g. elevation) to the use of a location by the animal."[25]: 745 For example, a habitat might be an aquatic or terrestrial environment that can be further categorized as a montane or alpine ecosystem. Habitat shifts provide important evidence of competition in nature where one population changes relative to the habitats that most other individuals of the species occupy. For example, one population of a species of tropical lizard (Tropidurus hispidus) has a flattened body relative to the main populations that live in open savanna. The population that lives in an isolated rock outcrop hides in crevasses where its flattened body offers a selective advantage. Habitat shifts also occur in the developmental life history of amphibians, and in insects that transition from aquatic to terrestrial habitats. Biotope and habitat are sometimes used interchangeably, but the former applies to a community's environment, whereas the latter applies to a species' environment.[24][26][27] Niche Main article: Ecological niche Termite mounds with varied heights of chimneys regulate gas exchange, temperature and other environmental parameters that are needed to sustain the internal physiology of the entire colony.[28][29] Definitions of the niche date back to 1917,[30] but G. Evelyn Hutchinson made conceptual advances in 1957[31][32] by introducing a widely adopted definition: "the set of biotic and abiotic conditions in which a species is able to persist and maintain stable population sizes."[30]: 519 The ecological niche is a central concept in the ecology of organisms and is sub-divided into the fundamental and the realized niche. The fundamental niche is the set of environmental conditions under which a species is able to persist. The realized niche is the set of environmental plus ecological conditions under which a species persists.[30][32][33] The Hutchinsonian niche is defined more technically as a "Euclidean hyperspace whose dimensions are defined as environmental variables and whose size is a function of the number of values that the environmental values may assume for which an organism has positive fitness."[34]: 71 Biogeographical patterns and range distributions are explained or predicted through knowledge of a species' traits and niche requirements.[35] Species have functional traits that are uniquely adapted to the ecological niche. A trait is a measurable property, phenotype, or characteristic of an organism that may influence its survival. Genes play an important role in the interplay of development and environmental expression of traits.[36] Resident species evolve traits that are fitted to the selection pressures of their local environment. This tends to afford them a competitive advantage and discourages similarly adapted species from having an overlapping geographic range. The competitive exclusion principle states that two species cannot coexist indefinitely by living off the same limiting resource; one will always out-compete the other. When similarly adapted species overlap geographically, closer inspection reveals subtle ecological differences in their habitat or dietary requirements.[37] Some models and empirical studies, however, suggest that disturbances can stabilize the co-evolution and shared niche occupancy of similar species inhabiting species-rich communities.[38] The habitat plus the niche is called the ecotope, which is defined as the full range of environmental and biological variables affecting an entire species.[24] Niche construction Main article: Niche construction See also: Ecosystem engineering Organisms are subject to environmental pressures, but they also modify their habitats. The regulatory feedback between organisms and their environment can affect conditions from local (e.g., a beaver pond) to global scales, over time and even after death, such as decaying logs or silica skeleton deposits from marine organisms.[39] The process and concept of ecosystem engineering are related to niche construction, but the former relates only to the physical modifications of the habitat whereas the latter also considers the evolutionary implications of physical changes to the environment and the feedback this causes on the process of natural selection. Ecosystem engineers are defined as: "organisms that directly or indirectly modulate the availability of resources to other species, by causing physical state changes in biotic or abiotic materials. In so doing they modify, maintain and create habitats."[40]: 373 The ecosystem engineering concept has stimulated a new appreciation for the influence that organisms have on the ecosystem and evolutionary process. The term "niche construction" is more often used in reference to the under-appreciated feedback mechanisms of natural selection imparting forces on the abiotic niche.[28][41] An example of natural selection through ecosystem engineering occurs in the nests of social insects, including ants, bees, wasps, and termites. There is an emergent homeostasis or homeorhesis in the structure of the nest that regulates, maintains and defends the physiology of the entire colony. Termite mounds, for example, maintain a constant internal temperature through the design of air-conditioning chimneys. The structure of the nests themselves is subject to the forces of natural selection. Moreover, a nest can survive over successive generations, so that progeny inherit both genetic material and a legacy niche that was constructed before their time.[5][28][29] Biome Main article: Biome Biomes are larger units of organization that categorize regions of the Earth's ecosystems, mainly according to the structure and composition of vegetation.[42] There are different methods to define the continental boundaries of biomes dominated by different functional types of vegetative communities that are limited in distribution by climate, precipitation, weather, and other environmental variables. Biomes include tropical rainforest, temperate broadleaf and mixed forest, temperate deciduous forest, taiga, tundra, hot desert, and polar desert.[43] Other researchers have recently categorized other biomes, such as the human and oceanic microbiomes. To a microbe, the human body is a habitat and a landscape.[44] Microbiomes were discovered largely through advances in molecular genetics, which have revealed a hidden richness of microbial diversity on the planet. The oceanic microbiome plays a significant role in the ecological biogeochemistry of the planet's oceans.[45] Biosphere Main article: Biosphere See also: Earth's spheres The largest scale of ecological organization is the biosphere: the total sum of ecosystems on the planet. Ecological relationships regulate the flux of energy, nutrients, and climate all the way up to the planetary scale. For example, the dynamic history of the planetary atmosphere's CO2 and O2 composition has been affected by the biogenic flux of gases coming from respiration and photosynthesis, with levels fluctuating over time in relation to the ecology and evolution of plants and animals.[46] Ecological theory has also been used to explain self-emergent regulatory phenomena at the planetary scale: for example, the Gaia hypothesis is an example of holism applied in ecological theory.[47] The Gaia hypothesis states that there is an emergent feedback loop generated by the metabolism of living organisms that maintains the core temperature of the Earth and atmospheric conditions within a narrow self-regulating range of tolerance.[48] Population ecology Main article: Population ecology See also: Lists of organisms by population Population ecology studies the dynamics of species populations and how these populations interact with the wider environment.[5] A population consists of individuals of the same species that live, interact, and migrate through the same niche and habitat.[49] A primary law of population ecology is the Malthusian growth model[50] which states, "a population will grow (or decline) exponentially as long as the environment experienced by all individuals in the population remains constant."[50]: 18 Simplified population models usually starts with four variables: death, birth, immigration, and emigration. An example of an introductory population model describes a closed population, such as on an island, where immigration and emigration does not take place. Hypotheses are evaluated with reference to a null hypothesis which states that random processes create the observed data. In these island models, the rate of population change is described by: 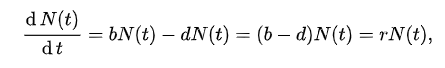 where N is the total number of individuals in the population, b and d are the per capita rates of birth and death respectively, and r is the per capita rate of population change.[50][51] Using these modeling techniques, Malthus' population principle of growth was later transformed into a model known as the logistic equation by Pierre Verhulst: 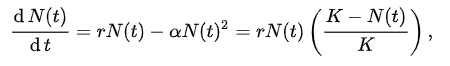  Metapopulations and migration Main article: Metapopulation See also: Animal migration The concept of metapopulations was defined in 1969[55] as "a population of populations which go extinct locally and recolonize".[56]: 105 Metapopulation ecology is another statistical approach that is often used in conservation research.[57] Metapopulation models simplify the landscape into patches of varying levels of quality,[58] and metapopulations are linked by the migratory behaviours of organisms. Animal migration is set apart from other kinds of movement because it involves the seasonal departure and return of individuals from a habitat.[59] Migration is also a population-level phenomenon, as with the migration routes followed by plants as they occupied northern post-glacial environments. Plant ecologists use pollen records that accumulate and stratify in wetlands to reconstruct the timing of plant migration and dispersal relative to historic and contemporary climates. These migration routes involved an expansion of the range as plant populations expanded from one area to another. There is a larger taxonomy of movement, such as commuting, foraging, territorial behavior, stasis, and ranging. Dispersal is usually distinguished from migration because it involves the one-way permanent movement of individuals from their birth population into another population.[60][61] In metapopulation terminology, migrating individuals are classed as emigrants (when they leave a region) or immigrants (when they enter a region), and sites are classed either as sources or sinks. A site is a generic term that refers to places where ecologists sample populations, such as ponds or defined sampling areas in a forest. Source patches are productive sites that generate a seasonal supply of juveniles that migrate to other patch locations. Sink patches are unproductive sites that only receive migrants; the population at the site will disappear unless rescued by an adjacent source patch or environmental conditions become more favorable. Metapopulation models examine patch dynamics over time to answer potential questions about spatial and demographic ecology. The ecology of metapopulations is a dynamic process of extinction and colonization. Small patches of lower quality (i.e., sinks) are maintained or rescued by a seasonal influx of new immigrants. A dynamic metapopulation structure evolves from year to year, where some patches are sinks in dry years and are sources when conditions are more favorable. Ecologists use a mixture of computer models and field studies to explain metapopulation structure.[62][63] Community ecology  Interspecific interactions such as predation are a key aspect of community ecology. Main article: Community ecology Community ecology examines how interactions among species and their environment affect the abundance, distribution and diversity of species within communities. Johnson & Stinchcomb (2007)[64]: 250 Community ecology is the study of the interactions among a collection of species that inhabit the same geographic area. Community ecologists study the determinants of patterns and processes for two or more interacting species. Research in community ecology might measure species diversity in grasslands in relation to soil fertility. It might also include the analysis of predator-prey dynamics, competition among similar plant species, or mutualistic interactions between crabs and corals. Ecosystem ecology Main article: Ecosystem ecology These ecosystems, as we may call them, are of the most various kinds and sizes. They form one category of the multitudinous physical systems of the universe, which range from the universe as a whole down to the atom. Tansley (1935)[65]: 299  A riparian forest in the White Mountains, New Hampshire (US) is an example of ecosystem ecology Ecosystems may be habitats within biomes that form an integrated whole and a dynamically responsive system having both physical and biological complexes. Ecosystem ecology is the science of determining the fluxes of materials (e.g. carbon, phosphorus) between different pools (e.g., tree biomass, soil organic material). Ecosystem ecologists attempt to determine the underlying causes of these fluxes. Research in ecosystem ecology might measure primary production (g C/m^2) in a wetland in relation to decomposition and consumption rates (g C/m^2/y). This requires an understanding of the community connections between plants (i.e., primary producers) and the decomposers (e.g., fungi and bacteria),[66] The underlying concept of an ecosystem can be traced back to 1864 in the published work of George Perkins Marsh ("Man and Nature").[67][68] Within an ecosystem, organisms are linked to the physical and biological components of their environment to which they are adapted.[65] Ecosystems are complex adaptive systems where the interaction of life processes form self-organizing patterns across different scales of time and space.[69] Ecosystems are broadly categorized as terrestrial, freshwater, atmospheric, or marine. Differences stem from the nature of the unique physical environments that shapes the biodiversity within each. A more recent addition to ecosystem ecology are technoecosystems, which are affected by or primarily the result of human activity.[5] Food webs Main article: Food web See also: Food chain 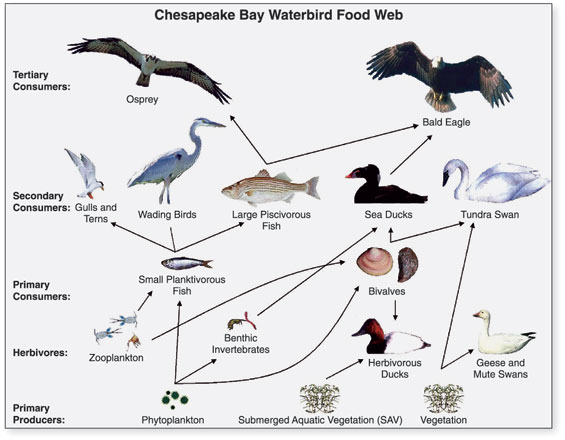 Generalized food web of waterbirds from Chesapeake Bay A food web is the archetypal ecological network. Plants capture solar energy and use it to synthesize simple sugars during photosynthesis. As plants grow, they accumulate nutrients and are eaten by grazing herbivores, and the energy is transferred through a chain of organisms by consumption. The simplified linear feeding pathways that move from a basal trophic species to a top consumer is called the food chain. Food chains in an ecological community create a complex food web. Food webs are a type of concept map that is used to illustrate and study pathways of energy and material flows.[7][70][71] Empirical measurements are generally restricted to a specific habitat, such as a cave or a pond, and principles gleaned from small-scale studies are extrapolated to larger systems.[72] Feeding relations require extensive investigations, e.g. into the gut contents of organisms, which can be difficult to decipher, or stable isotopes can be used to trace the flow of nutrient diets and energy through a food web.[73] Despite these limitations, food webs remain a valuable tool in understanding community ecosystems.[74] Food webs illustrate important principles of ecology: some species have many weak feeding links (e.g., omnivores) while some are more specialized with fewer stronger feeding links (e.g., primary predators). Such linkages explain how ecological communities remain stable over time[75][76] and eventually can illustrate a "complete" web of life.[71][77][78][79] The disruption of food webs may have a dramatic impact on the ecology of individual species or whole ecosystems. For instance, the replacement of an ant species by another (invasive) ant species has been shown to affect how elephants reduce tree cover and thus the predation of lions on zebras.[80][81] Trophic levels Main article: Trophic level 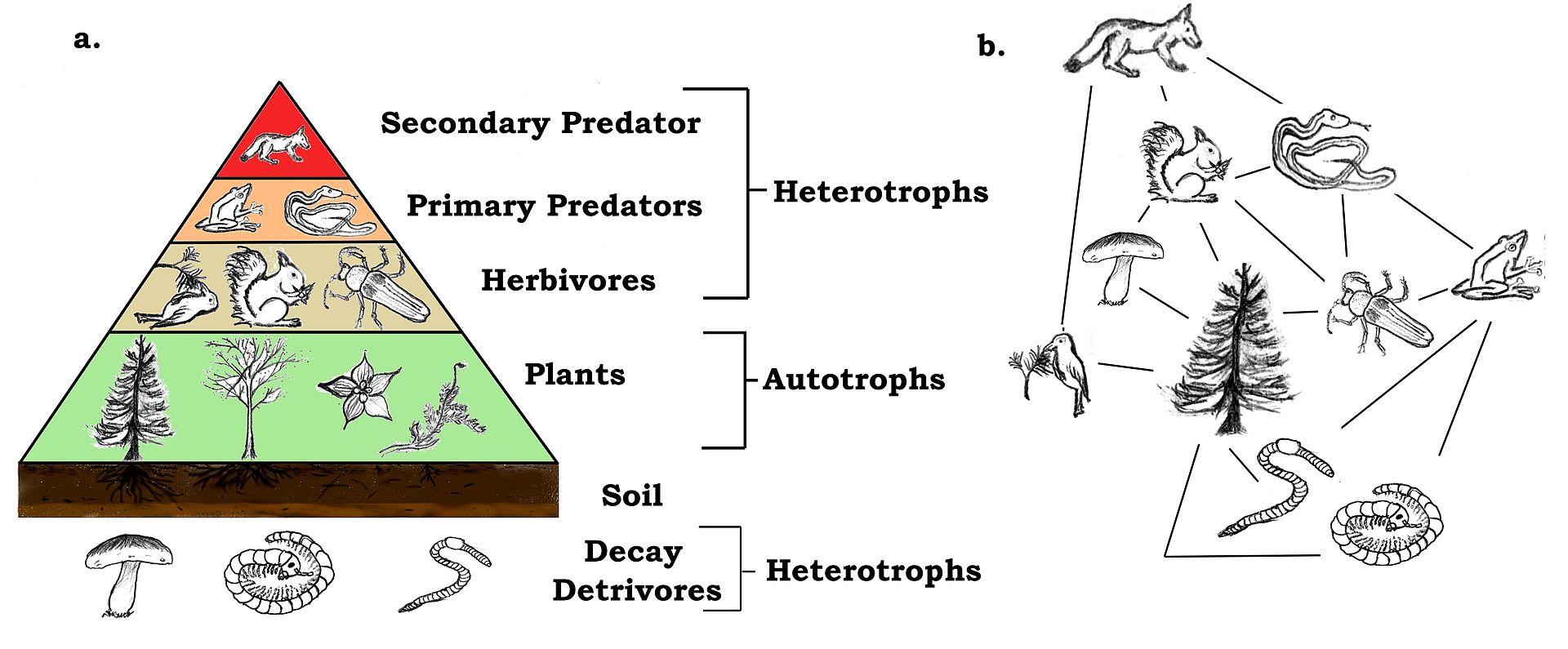 A trophic pyramid (a) and a food-web (b) illustrating ecological relationships among creatures that are typical of a northern boreal terrestrial ecosystem. The trophic pyramid roughly represents the biomass (usually measured as total dry-weight) at each level. Plants generally have the greatest biomass. Names of trophic categories are shown to the right of the pyramid. Some ecosystems, such as many wetlands, do not organize as a strict pyramid, because aquatic plants are not as productive as long-lived terrestrial plants such as trees. Ecological trophic pyramids are typically one of three kinds: 1) pyramid of numbers, 2) pyramid of biomass, or 3) pyramid of energy.[5]: 598 A trophic level (from Greek troph, τροφή, trophē, meaning "food" or "feeding") is "a group of organisms acquiring a considerable majority of its energy from the lower adjacent level (according to ecological pyramids) nearer the abiotic source."[82]: 383 Links in food webs primarily connect feeding relations or trophism among species. Biodiversity within ecosystems can be organized into trophic pyramids, in which the vertical dimension represents feeding relations that become further removed from the base of the food chain up toward top predators, and the horizontal dimension represents the abundance or biomass at each level.[83] When the relative abundance or biomass of each species is sorted into its respective trophic level, they naturally sort into a 'pyramid of numbers'.[84] Species are broadly categorized as autotrophs (or primary producers), heterotrophs (or consumers), and Detritivores (or decomposers). Autotrophs are organisms that produce their own food (production is greater than respiration) by photosynthesis or chemosynthesis. Heterotrophs are organisms that must feed on others for nourishment and energy (respiration exceeds production).[5] Heterotrophs can be further sub-divided into different functional groups, including primary consumers (strict herbivores), secondary consumers (carnivorous predators that feed exclusively on herbivores), and tertiary consumers (predators that feed on a mix of herbivores and predators).[85] Omnivores do not fit neatly into a functional category because they eat both plant and animal tissues. It has been suggested that omnivores have a greater functional influence as predators because compared to herbivores, they are relatively inefficient at grazing.[86] Trophic levels are part of the holistic or complex systems view of ecosystems.[87][88] Each trophic level contains unrelated species that are grouped together because they share common ecological functions, giving a macroscopic view of the system.[89] While the notion of trophic levels provides insight into energy flow and top-down control within food webs, it is troubled by the prevalence of omnivory in real ecosystems. This has led some ecologists to "reiterate that the notion that species clearly aggregate into discrete, homogeneous trophic levels is fiction."[90]: 815 Nonetheless, recent studies have shown that real trophic levels do exist, but "above the herbivore trophic level, food webs are better characterized as a tangled web of omnivores."[91]: 612 Keystone species Main article: Keystone species  Sea otters, an example of a keystone species A keystone species is a species that is connected to a disproportionately large number of other species in the food-web. Keystone species have lower levels of biomass in the trophic pyramid relative to the importance of their role. The many connections that a keystone species holds means that it maintains the organization and structure of entire communities. The loss of a keystone species results in a range of dramatic cascading effects (termed trophic cascades) that alters trophic dynamics, other food web connections, and can cause the extinction of other species.[92][93] The term keystone species was coined by Robert Paine in 1969 and is a reference to the keystone architectural feature as the removal of a keystone species can result in a community collapse just as the removal of the keystone in an arch can result in the arch's loss of stability.[94] Sea otters (Enhydra lutris) are commonly cited as an example of a keystone species because they limit the density of sea urchins that feed on kelp. If sea otters are removed from the system, the urchins graze until the kelp beds disappear, and this has a dramatic effect on community structure.[95] Hunting of sea otters, for example, is thought to have led indirectly to the extinction of the Steller's sea cow (Hydrodamalis gigas).[96] While the keystone species concept has been used extensively as a conservation tool, it has been criticized for being poorly defined from an operational stance. It is difficult to experimentally determine what species may hold a keystone role in each ecosystem. Furthermore, food web theory suggests that keystone species may not be common, so it is unclear how generally the keystone species model can be applied.[95][97] |
組織のレベル、範囲、規模 主な記事 生態学の概要 生態学の範囲には、ミクロレベル(細胞など)から惑星スケール(生物圏など)の現象まで、相互作用するさまざまなレベルの組織が含まれる。例えば、生態系 は、生物資源と相互作用する生命体(すなわち、個体群に集合する個々の生物)を含む。生態系は動的であり、必ずしも直線的な遷移ルートをたどるわけではな いため、特定の森林遷移段階が生物学的プロセスによってもたらされるまでに、数千年かけて急速に、あるいはゆっくりと変化が起こるかもしれない。生態系の 面積は、小さなものから広大なものまで実に様々である。1本の木は森林生態系の分類にはほとんど関係ないが、その中や上に生息する生物にとっては決定的に 重要である[2]。生態学的群集におけるつながりの性質は、それぞれの種の詳細を単独で知ることでは説明できない。なぜなら、生態系が統合された全体とし て研究されるまで、創発的なパターンは明らかにならず、予測もできないからである。 生態学の主な下位分野である個体群生態学(または群集生態学)と生態系生態学は、規模の違いだけでなく、この分野における2つの対照的なパラダイムの違いを示している。前者は生物の分布と存在量に焦点を当て、後者は物質とエネルギーのフラックスに焦点を当てている[6]。 階層 以下も参照のこと: 生物組織と生物分類 システムの行動は、まず組織のさまざまなレベルに分類されなければならない。より高いレベルに対応する行動は、ゆっくりとした速度で起こる。逆に、組織レ ベルの低いものは、速い速度で起こる。例えば、個々の木の葉は、光強度やCO2濃度などの瞬間的な変化に素早く反応する。樹木の成長はよりゆっくりと反応 し、これらの短期的変化を統合する。 O'Neillら(1986)[7]: 76 生態系のダイナミクスのスケールは、アブラムシが1本の樹木上を移動するような閉じたシステムのように動作する一方で、大気や気候のようなより広いスケー ルの影響については開放的であり続けることができる。したがって生態学者は、植生群、気候、土壌の種類など、より細かいスケールの単位から収集したデータ を分析することによって生態系を階層的に分類し、これらの情報を統合して、局所的、地域的、景観的、年代的スケールで作用する均一な組織とプロセスの出現 パターンを特定する。 生態学の研究を概念的に管理可能な枠組みに構造化するため、生物界は、遺伝子から細胞、組織、器官、生物、種、個体群、ギルド、群集、生態系、生物圏、そ して生物圏のレベルまで、入れ子のような階層構造で構成されている。 [この枠組みはパナーキーを形成し[9]、非線形の挙動を示す。これは、「効果と原因が比例しないため、窒素固定因子の数などの重要な変数に対する小さな 変化が、システムの特性に比例しない、おそらく不可逆的な変化をもたらす」ことを意味する[10]: 14 生物多様性 主な記事 生物多様性 生物多様性とは、生命とそのプロセスの多様性を指す。生物多様性とは、生物の多様性、生物間の遺伝的差異、それらが存在する群集や生態系、そしてそれらを機能させながら常に変化し適応させている生態学的・進化的プロセスを指す。 Noss & Carpenter (1994)[11]による: 5 生物多様性(「生物学的多様性」の略)とは、遺伝子から生態系に至るまで、生物組織のあらゆるレベルにまたがる生命の多様性を表す言葉である。生物多様性 には、種の多様性、生態系の多様性、遺伝子の多様性が含まれ、科学者は、この多様性が、それぞれのレベルで、またこれらのレベル間で作用する複雑な生態学 的プロセスにどのような影響を与えるかに関心を持っている[12][13][14]。 [13][15][16]生物多様性は、人間の生活の質を維持・向上させる生態系サービスにおいて重要な役割を果たしている[14][17][18]。個 体群を支える自然資本は、生態系サービスを維持するために不可欠であり[19][20]、種の移動(例えば、河川における魚の遡上や鳥類の害虫駆除)は、 これらのサービスの損失が発生するメカニズムの1つとして関与している[21]。 生息地 主な記事 生息地 サンゴ礁の生物多様性。サンゴは炭酸カルシウムの骨格を形成することで、環境に適応し、環境を変化させる。これは、将来の世代に生育条件を提供し、他の多くの種の生息地を形成する[23]。 巣を作るオナガガモ ある種の生息地は、その種が生息することが知られている環境と、その結果として形成される群落のタイプを記述する[24] 。 例えば、生息地は水生環境または陸生環境であり、さらに山地生態系または高山生態系に分類することができる。生息域の移動は、ある個体群がその種の他のほ とんどの個体が占める生息域と相対的に変化する、自然界における競争の重要な証拠となる。例えば、ある種の熱帯トカゲ(Tropidurus hispidus)のある個体群は、開けたサバンナに生息する主な個体群と比べて、体が平らになっている。孤立した岩場に生息する個体群は、その扁平な体 が選択的に有利となるクレバスに身を隠す。生息地のシフトは両生類の発生生活史や、水生生息地から陸上生息地へ移行する昆虫でも起こる。ビオトープと生息 域は同じ意味で使われることもあるが、前者が生物群集の環境に適用されるのに対し、後者は種の環境に適用される[24][26][27]。 ニッチ 主な記事 生態学的ニッチ 様々な高さの煙突を持つシロアリ塚は、コロニー全体の内部生理を維持するために必要なガス交換、温度、その他の環境パラメータを調節している[28][29]。 ニッチの定義は1917年まで遡るが[30]、G. Evelyn Hutchinsonは1957年に広く採用されている定義を導入することで、概念的な進歩を遂げた[31][32]: 「ある種が存続し、安定した個体数を維持することができる生物学的・生物学的条件の集合」[30]である: 519 生態学的ニッチは生物の生態学における中心的な概念であり、基本的ニッチと実現ニッチに細分化される。基本ニッチとは、ある種が存続できる環境条件の集合 である。実現ニッチとは、種が存続する環境条件+生態学的条件の集合である[30][32][33]。ハッチンソン式ニッチは、より専門的には「次元が環 境変数として定義され、その大きさが、生物が正の適合度を持つ環境値がとりうる値の数の関数であるユークリッド超空間」と定義される[34]: 71。 生物地理学的パターンと生息域分布は、種の形質とニッチ要件に関する知識によって説明または予測される[35]。形質とは、生物の生存に影響を及ぼす可能 性のある測定可能な性質、表現型、特性のことである。遺伝子は、形質の発生と環境的発現の相互作用において重要な役割を果たす。このことは、それらの種に 競争上の優位性を与え、同様に適応した種が地理的に重複した範囲を持つことを抑制する傾向がある。競争的排除の原理は、2つの種が同じ制限資源を利用して 無限に共存することは不可能であるとしている。同じように適応した種が地理的に重複している場合、より詳細に調査すると、生息地や食餌要求における微妙な 生態学的差異が明らかになる[37]。しかし、いくつかのモデルや実証的研究は、撹乱が、種が豊富な群落に生息する類似種の共進化や共有ニッチ占有を安定 化させる可能性があることを示唆している[38]。生息地+ニッチはエコトープと呼ばれ、種全体に影響を与える環境・生物学的変数の全範囲として定義され る[24]。 ニッチの構築 主な記事 ニッチ構築 以下も参照のこと: 生態系工学 生物は環境からの圧力を受けるが、同時に生息環境をも変化させる。生物とその環境との間の調節フィードバックは、局所的なもの(例えばビーバーの池)から 地球規模のものまで、時間をかけて、さらには朽ちた丸太や海洋生物のシリカ骨格の堆積物のような死後の状態にまで影響を及ぼすことがある[39]。生態系 工学のプロセスと概念はニッチ構築と関連しているが、前者が生息地の物理的な改変のみに関連しているのに対し、後者は環境への物理的な変化とこれが自然淘 汰のプロセスに引き起こすフィードバックの進化的な意味合いも考慮している。生態系エンジニアは次のように定義される: 生物学的または生物学的物質に物理的な状態変化を引き起こすことによって、他の種が利用できる資源を直接的または間接的に調節する生物」と定義される。そ うすることで、生息地を変更、維持、創造する」[40]: 373 生態系工学の概念は、生物が生態系と進化のプロセスに与える影響に対する新たな認識を刺激した。ニッチ構築」という用語は、自然淘汰のフィードバック・メ カニズムが生物学的ニッチに与える力について、あまり評価されていないことを指して使われることが多くなっている[28][41]。生態系工学による自然 淘汰の例は、アリ、ハチ、スズメバチ、シロアリなどの社会性昆虫の巣に見られる。巣の構造には、コロニー全体の生理機能を制御し、維持し、防御する、創発 的なホメオスタシス(恒常性)が存在する。例えばシロアリの塚は、空調用の煙突の設計によって内部温度を一定に保っている。巣の構造そのものは、自然淘汰 の力を受けている。さらに、巣は何世代にもわたって存続することができるため、子孫は遺伝物質と、自分たちの時代以前に構築されたニッチを受け継ぐことに なる[5][28][29]。 バイオーム 主な記事 バイオーム バイオームとは、主に植生の構造と構成に従って、地球の生態系の地域を分類する大きな組織単位である[42]。気候、降水量、天候、その他の環境変数に よって分布が制限される、異なる機能的タイプの植生群落が支配するバイオームの大陸境界を定義する方法は様々である。バイオームには、熱帯雨林、温帯広葉 樹林と混交林、温帯落葉樹林、タイガ、ツンドラ、高温砂漠、極地砂漠などがある[43]。他の研究者は最近、ヒトや海洋のマイクロバイオームなど、他のバ イオームを分類している。微生物にとって、人体は一つの生息地であり、一つの景観である[44]。マイクロバイオームは、主に分子遺伝学の進歩によって発 見され、地球上に隠された微生物の多様性の豊かさを明らかにした。海洋マイクロバイオームは、地球上の海洋の生態生物地球化学において重要な役割を果たし ている[45]。 生物圏 主な記事 生物圏 以下も参照のこと: 地球の球体 生態系組織の最大のスケールは生物圏であり、地球上の生態系の総体である。生態学的関係は、エネルギー、栄養素、気候の流束を、惑星スケールまで制御して いる。例えば、惑星大気のCO2とO2組成の動的な歴史は、呼吸と光合成からもたらされるガスの生物起源フラックスによって影響を受けており、そのレベル は動植物の生態系と進化に関連して時間と共に変動している。 [例えば、ガイア仮説は、生態学理論に適用された全体論の一例である。ガイア仮説は、地球の中心温度と大気の状態を、自己調節可能な狭い許容範囲内に維持 する、生物の代謝によって生成される創発的なフィードバックループが存在すると述べている[48]。 個体群生態学 主な記事 個体群生態学 以下も参照のこと: 個体群別の生物リスト 個体群生態学は、種の個体群の動態と、これらの個体群がより広い環境とどのように相互作用するかを研究する学問である。 個体群生態学の主要な法則はマルサス成長モデルであり[50]、「個体群内のすべての個体が経験する環境が一定である限り、個体群は指数関数的に成長する (または減少する)」と述べている[50]: 18 簡略化された人口モデルは通常、死亡、出生、移民、移住という4つの変数から始まる。 入門的な個体群モデルの例としては、移民や移住が行われない、島のような閉鎖的な個体群を説明する。仮説は帰無仮説を参照して評価される。帰無仮説とは、 ランダムな過程が観察されたデータを作り出しているというものである。このような島嶼モデルでは、人口の変化率は次式で表される: ここで、Nは人口の総個体数、bとdはそれぞれ一人当たりの出生率と死亡率、rは一人当たりの人口変化率である[50][51]。 これらのモデリング技術を用いて、マルサスの人口増加原理は、後にピエール・ヴェルフルストによってロジスティック方程式として知られるモデルに変換された: メタ集団と移動 主な記事 メタポピュレーション も参照のこと: 動物の移動 メタ個体群の概念は1969年に定義された[55]。「局所的に絶滅し、再殖する個体群の集団」である[56]: 105 メタ個体群生態学は、保全研究でしばしば用いられるもう一つの統計的アプローチである[57]。メタ個体群モデルは、景観を様々なレベルの質のパッチに単 純化し[58]、メタ個体群は生物の移動行動によって結びつけられている。動物の移動は、生息地からの個体の季節的な出発と帰還を伴うため、他の種類の移 動とは区別される[59]。植物が氷河期後の北方環境を占領する際にたどった移動ルートのように、移動は個体群レベルの現象でもある。植物生態学者は、湿 地帯に蓄積・成層する花粉の記録を用いて、歴史的・現代的な気候に対する植物の移動・分散の時期を復元している。これらの移動ルートは、植物個体群がある 地域から別の地域へと拡大するにつれて、生息域の拡大を伴うものである。移動には、通勤、採食、縄張り行動、静止、放散などの大きな分類法がある。分散は 通常、出生集団から別の集団への個体の一方向の永続的な移動を伴うため、移動とは区別される[60][61]。 メタ個体群の用語では、移動する個体は移住(ある地域から出る場合)または移民(ある地域に入る場合)に分類され、サイトは発生源または吸収源に分類され る。サイトとは、生態学者が個体群をサンプリングする場所の総称で、池や森林内の定義されたサンプリング・エリアなどを指す。ソース・パッチは生産性の高 い場所で、他のパッチに移動する幼生を季節的に供給する。シンクパッチとは、移動のみを受け入れる非生産的な場所のことで、隣接するソースパッチによって 救出されるか、環境条件がより良好にならない限り、その場所の個体群は消滅する。メタ個体群モデルは、空間生態学や個体群統計学に関する潜在的な疑問に答 えるために、経時的なパッチの動態を調べる。メタ個体群の生態は、絶滅とコロニー化のダイナミックなプロセスである。質の低い小さなパッチ(すなわちシン ク)は、季節的な新しい移民の流入によって維持されたり、救済されたりする。ダイナミックなメタ個体群構造は年ごとに進化し、乾燥した年にはいくつかの パッチがシンクになり、条件が良くなるとソースになる。生態学者は、コンピュータ・モデルと野外調査を組み合わせて、メタ個体群構造を説明している [62][63]。 群集生態学 捕食などの種間相互作用は、群集生態学の重要な側面である。 主な記事 群集生態学 群集生態学では、種とその環境との相互作用が、群集内の種の存在量、分布、多様性にどのような影響を与えるかを調べる。 Johnson & Stinchcomb (2007)[64]: 250 群集生態学は、同じ地域に生息する種の集合体間の相互作用を研究する学問である。群集生態学者は、相互作用する2つ以上の種のパターンとプロセスの決定要 因を研究する。群集生態学の研究では、土壌肥沃度との関係で草原における種の多様性を測定することもある。また、捕食者と被食者の動態、類似植物種間の競 争、カニとサンゴの相互作用の分析も含まれる。 生態系生態学 主な記事 生態系生態学 生態系と呼ばれるものには、さまざまな種類と規模がある。それらは宇宙全体から原子に至るまで、宇宙の膨大な物理システムのひとつのカテゴリーを形成している。 タンズレー (1935)[65]: 299 ニューハンプシャー州ホワイトマウンテンズ(米国)の水辺林は、生態系生態学の一例である。 生態系は、物理的および生物学的複合体の両方を有する統合された全体および動的に応答するシステムを形成する生物群内の生息地である。生態系生態学は、異 なるプール(樹木バイオマス、土壌有機物など)間の物質(炭素、リンなど)のフラックスを決定する科学である。生態系生態学者は、これらのフラックスの根 本的な原因を特定しようと試みる。生態系生態学の研究では、湿地の一次生産量(g C/m^2)を分解・消費速度(g C/m^2/y)との関係で測定することがある。これには、植物(すなわち、一次生産者)と分解者(例えば、菌類やバクテリア)の間の群集のつながりを理 解する必要がある[66]。 生態系の根底にある概念は、1864年にジョージ・パーキンス・マーシュが発表した著作(『人間と自然』)にまで遡ることができる[67][68]。生態 系の中では、生物は、それらが適応している環境の物理的・生物学的構成要素と結びついている[65]。生態系は、生命プロセスの相互作用が、時間と空間の さまざまなスケールにわたって自己組織化パターンを形成する複雑な適応システムである[69]。生態系は、陸上、淡水、大気、海洋に大別される。その違い は、それぞれの生物多様性を形成する独自の物理的環境の性質に起因する。生態系生態学に最近追加されたものにテクノ生態系がある。テクノ生態系は、人間活 動の影響を受けたり、主に人間活動の結果である。 食物網 主な記事 食物網 以下も参照のこと: 食物連鎖 チェサピーク湾の水鳥の一般的な食物網 食物網は典型的な生態系ネットワークである。植物は太陽エネルギーを取り込み、それを使って光合成を行い、単糖を合成する。植物は成長するにつれて栄養分 を蓄積し、草を食む草食動物に食べられ、そのエネルギーは消費されることによって生物の連鎖を通じて伝達される。基礎栄養種から上位消費者へと移動する単 純化された直線的な摂食経路を食物連鎖と呼ぶ。生態系群集における食物連鎖は、複雑な食物網を形成する。食物網は、エネルギーと物質の流れの経路を図示 し、研究するために使用される概念図の一種である[7][70][71]。 経験的な測定は、一般的に洞窟や池などの特定の生息域に限定され、小規模な研究から得られた原則は、より大規模な系に外挿される[72]。摂食関係は、例 えば、解読が困難な生物の腸内容物への広範な調査を必要とするか、または食物網を介した栄養飼料とエネルギーの流れを追跡するために安定同位体を使用する ことができる[73]。これらの制限にもかかわらず、食物網は、依然として群集生態系を理解するための貴重なツールである[74]。 食物網は生態学の重要な原則を示している。ある種は多くの弱い摂食リンクを持ち(例:雑食動物)、ある種はより強い摂食リンクが少なく、より特化している (例:一次捕食者)。このような連鎖は、生態学的群集が時間とともに安定した状態を保ち[75][76]、最終的には「完全な」生命の網を示すことができ ることを説明している[71][77][78][79]。 食物網の崩壊は、個々の種や生態系全体の生態系に劇的な影響を与える可能性がある。例えば、あるアリ種が別の(侵略的な)アリ種に置き換わることで、ゾウが樹木の被覆を減らし、その結果ライオンがシマウマを捕食する方法に影響を与えることが示されている[80][81]。 栄養段階 主な記事 栄養段階 栄養ピラミッド(a)と食物網(b)は、北方寒帯の陸上生態系に典型的な生物間の生態学的関係を示している。栄養ピラミッドは、各レベルのバイオマス(通 常、総乾燥重量で測定)を大まかに表している。一般に、植物が最も大きなバイオマスを持つ。ピラミッドの右側には、栄養区分の名前が示されている。水生植 物は、樹木のような長命な陸上植物ほど生産性が高くないため、多くの湿地帯のように、厳密なピラミッド型にならない生態系もある。生態系の栄養ピラミッド は、通常、1)数のピラミッド、2)バイオマスのピラミッド、3)エネルギーのピラミッドの3種類のうちの1つである[5]: 598 栄養段階(ギリシャ語のtroph、τροφή、trophēに由来し、「食物」または「摂食」を意味する)とは、「(生態ピラミッドによれば)生物源に 近い下位の隣接するレベルから、そのエネルギーのかなりの大部分を獲得する生物群」のことである[82]: 383 食物網のリンクは、主に種間の摂食関係や栄養主義をつなぐものである。生態系内の生物多様性は、栄養ピラミッドに整理することができる。このピラミッドで は、縦の次元が食物連鎖の底辺から上位の捕食者に向かってさらに離れていく摂食関係を表し、横の次元が各レベルにおける存在量またはバイオマスを表してい る[83]。各生物種の相対的な存在量またはバイオマスをそれぞれの栄養レベルに分類すると、それらは自然に「数のピラミッド」に分類される[84]。 種は大きく、独立栄養生物(または一次生産者)、従属栄養生物(または消費者)、および腐食生物(または分解者)に分類される。独立栄養生物は、光合成や 化学合成によって自ら食物を生産する(生産量が呼吸量を上回る)生物である。従属栄養生物は、栄養とエネルギーを得るために他者を捕食しなければならない (呼吸が生産を上回る)生物である。従属栄養生物はさらに、一次消費者(厳密な草食動物)、二次消費者(草食動物のみを捕食する肉食捕食動物)、三次消費 者(草食動物と捕食動物の混合物を捕食する捕食動物)など、さまざまな機能グループに細分化できる。雑食動物は草食動物に比べ、放牧の効率が比較的悪いた め、捕食者としての機能的影響力が大きいことが示唆されている[86]。 栄養段階は、生態系を全体的または複雑系的にとらえる考え方の一部である[87][88]。各栄養段階には、生態学的機能を共有しているためにグループ化 された無関係な種が含まれ、生態系を巨視的にとらえることができる[89]。栄養段階という概念は、食物網内のエネルギーの流れやトップダウン制御に関す る洞察を与える一方で、実際の生態系では雑食が多いという問題を抱えている。このため、一部の生態学者は、「種が明らかに離散的で均質な栄養レベルに集約 されるという考え方は虚構であると繰り返し述べている」[90]: 815にもかかわらず、最近の研究では、実際の栄養レベルは存在するが、「草食動物の栄養レベル以上では、食物網は雑食動物のもつれた網としてよりよく特 徴づけられる」ことが示されている[91]: 612。 要となる種 主な記事 要となる種 ラッコ、キーストーン種の一例 キーストーン種とは、食物網の中で不釣り合いに多くの他の種とつながっている種のことである。キーストーン種は、その役割の重要性に比べて、栄養ピラミッ ドにおけるバイオマスのレベルが低い。要となる種が持つ多くのつながりは、その種が生物群集全体の組織と構造を維持することを意味する。キーストーン種の 喪失は、栄養動態や他の食物網のつながりを変化させる様々な劇的な連鎖的影響(栄養カスケードと呼ばれる)をもたらし、他の種の絶滅を引き起こす可能性が ある[92][93]。キーストーン種という用語は、1969年にロバート・ペインによって作られた造語であり、アーチのキーストーンが除去されるとアー チの安定性が失われるように、キーストーン種の除去は群集の崩壊をもたらす可能性があるとして、キーストーン建築の特徴に言及したものである[94]。 ラッコ(Enhydra lutris)は、ケルプを餌とするウニの密度を制限するため、一般的にキーストーン種の例として挙げられる。例えば、ラッコの狩猟は、間接的にトドウミ ウシ(Hydrodamalis gigas)の絶滅につながったと考えられている[96]。キーストーン種の概念は、保全ツールとして広く用いられてきたが、運用上の定義が不十分である という批判もある。それぞれの生態系において、どのような種が要となりうるかを実験的に決定することは困難である。さらに、食物網理論では、要となる種は 一般的ではない可能性が示唆されているため、要となる種モデルをどの程度一般的に適用できるかは不明である[95][97]。 |
| Complexity Main article: Complexity See also: Emergence Complexity is understood as a large computational effort needed to piece together numerous interacting parts exceeding the iterative memory capacity of the human mind. Global patterns of biological diversity are complex. This biocomplexity stems from the interplay among ecological processes that operate and influence patterns at different scales that grade into each other, such as transitional areas or ecotones spanning landscapes. Complexity stems from the interplay among levels of biological organization as energy, and matter is integrated into larger units that superimpose onto the smaller parts. "What were wholes on one level become parts on a higher one."[98]: 209 Small scale patterns do not necessarily explain large scale phenomena, otherwise captured in the expression (coined by Aristotle) 'the sum is greater than the parts'.[99][100][E] "Complexity in ecology is of at least six distinct types: spatial, temporal, structural, process, behavioral, and geometric."[101]: 3 From these principles, ecologists have identified emergent and self-organizing phenomena that operate at different environmental scales of influence, ranging from molecular to planetary, and these require different explanations at each integrative level.[48][102] Ecological complexity relates to the dynamic resilience of ecosystems that transition to multiple shifting steady-states directed by random fluctuations of history.[9][103] Long-term ecological studies provide important track records to better understand the complexity and resilience of ecosystems over longer temporal and broader spatial scales. These studies are managed by the International Long Term Ecological Network (LTER).[104] The longest experiment in existence is the Park Grass Experiment, which was initiated in 1856.[105] Another example is the Hubbard Brook study, which has been in operation since 1960.[106] Holism Main article: Holism Holism remains a critical part of the theoretical foundation in contemporary ecological studies. Holism addresses the biological organization of life that self-organizes into layers of emergent whole systems that function according to non-reducible properties. This means that higher-order patterns of a whole functional system, such as an ecosystem, cannot be predicted or understood by a simple summation of the parts.[107] "New properties emerge because the components interact, not because the basic nature of the components is changed."[5]: 8 Ecological studies are necessarily holistic as opposed to reductionistic.[36][102][108] Holism has three scientific meanings or uses that identify with ecology: 1) the mechanistic complexity of ecosystems, 2) the practical description of patterns in quantitative reductionist terms where correlations may be identified but nothing is understood about the causal relations without reference to the whole system, which leads to 3) a metaphysical hierarchy whereby the causal relations of larger systems are understood without reference to the smaller parts. Scientific holism differs from mysticism that has appropriated the same term. An example of metaphysical holism is identified in the trend of increased exterior thickness in shells of different species. The reason for a thickness increase can be understood through reference to principles of natural selection via predation without the need to reference or understand the biomolecular properties of the exterior shells.[109] +++++++++++++++++++++++++++++++++++++++++++ Relation to evolution Main article: Evolutionary ecology Ecology and evolutionary biology are considered sister disciplines of the life sciences. Natural selection, life history, development, adaptation, populations, and inheritance are examples of concepts that thread equally into ecological and evolutionary theory. Morphological, behavioural, and genetic traits, for example, can be mapped onto evolutionary trees to study the historical development of a species in relation to their functions and roles in different ecological circumstances. In this framework, the analytical tools of ecologists and evolutionists overlap as they organize, classify, and investigate life through common systematic principles, such as phylogenetics or the Linnaean system of taxonomy.[110] The two disciplines often appear together, such as in the title of the journal Trends in Ecology and Evolution.[111] There is no sharp boundary separating ecology from evolution, and they differ more in their areas of applied focus. Both disciplines discover and explain emergent and unique properties and processes operating across different spatial or temporal scales of organization.[36][48] While the boundary between ecology and evolution is not always clear, ecologists study the abiotic and biotic factors that influence evolutionary processes,[112][113] and evolution can be rapid, occurring on ecological timescales as short as one generation.[114] Behavioural ecology Main article: Behavioural ecology 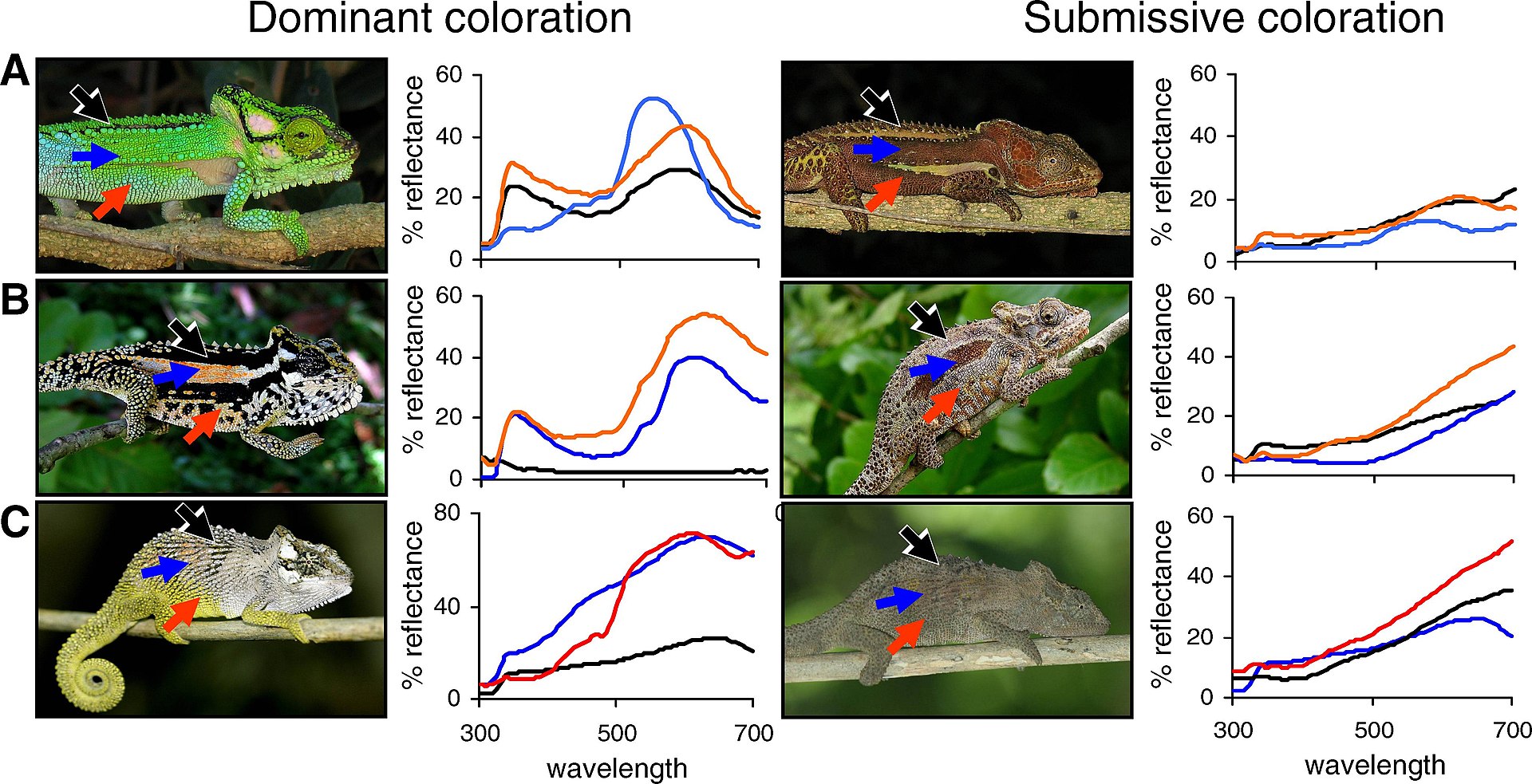 Social display and colour variation in differently adapted species of chameleons (Bradypodion spp.). Chameleons change their skin colour to match their background as a behavioural defence mechanism and also use colour to communicate with other members of their species, such as dominant (left) versus submissive (right) patterns shown in the three species (A-C) above.[115] All organisms can exhibit behaviours. Even plants express complex behaviour, including memory and communication.[116] Behavioural ecology is the study of an organism's behaviour in its environment and its ecological and evolutionary implications. Ethology is the study of observable movement or behaviour in animals. This could include investigations of motile sperm of plants, mobile phytoplankton, zooplankton swimming toward the female egg, the cultivation of fungi by weevils, the mating dance of a salamander, or social gatherings of amoeba.[117][118][119][120][121] Adaptation is the central unifying concept in behavioural ecology.[122] Behaviours can be recorded as traits and inherited in much the same way that eye and hair colour can. Behaviours can evolve by means of natural selection as adaptive traits conferring functional utilities that increases reproductive fitness.[123][124]  Mutualism: Leafhoppers (Eurymela fenestrata) are protected by ants (Iridomyrmex purpureus) in a mutualistic relationship. The ants protect the leafhoppers from predators and stimulate feeding in the leafhoppers, and in return, the leafhoppers feeding on plants exude honeydew from their anus that provides energy and nutrients to tending ants.[125] Predator-prey interactions are an introductory concept into food-web studies as well as behavioural ecology.[126] Prey species can exhibit different kinds of behavioural adaptations to predators, such as avoid, flee, or defend. Many prey species are faced with multiple predators that differ in the degree of danger posed. To be adapted to their environment and face predatory threats, organisms must balance their energy budgets as they invest in different aspects of their life history, such as growth, feeding, mating, socializing, or modifying their habitat. Hypotheses posited in behavioural ecology are generally based on adaptive principles of conservation, optimization, or efficiency.[33][112][127] For example, "[t]he threat-sensitive predator avoidance hypothesis predicts that prey should assess the degree of threat posed by different predators and match their behaviour according to current levels of risk"[128] or "[t]he optimal flight initiation distance occurs where expected postencounter fitness is maximized, which depends on the prey's initial fitness, benefits obtainable by not fleeing, energetic escape costs, and expected fitness loss due to predation risk."[129] Elaborate sexual displays and posturing are encountered in the behavioural ecology of animals. The birds-of-paradise, for example, sing and display elaborate ornaments during courtship. These displays serve a dual purpose of signalling healthy or well-adapted individuals and desirable genes. The displays are driven by sexual selection as an advertisement of quality of traits among suitors.[130] Cognitive ecology Cognitive ecology integrates theory and observations from evolutionary ecology and neurobiology, primarily cognitive science, in order to understand the effect that animal interaction with their habitat has on their cognitive systems and how those systems restrict behavior within an ecological and evolutionary framework.[131] "Until recently, however, cognitive scientists have not paid sufficient attention to the fundamental fact that cognitive traits evolved under particular natural settings. With consideration of the selection pressure on cognition, cognitive ecology can contribute intellectual coherence to the multidisciplinary study of cognition."[132][133] As a study involving the 'coupling' or interactions between organism and environment, cognitive ecology is closely related to enactivism,[131] a field based upon the view that "...we must see the organism and environment as bound together in reciprocal specification and selection...".[134] Social ecology Main article: Social ecology (academic field) Social-ecological behaviours are notable in the social insects, slime moulds, social spiders, human society, and naked mole-rats where eusocialism has evolved. Social behaviours include reciprocally beneficial behaviours among kin and nest mates[119][124][135] and evolve from kin and group selection. Kin selection explains altruism through genetic relationships, whereby an altruistic behaviour leading to death is rewarded by the survival of genetic copies distributed among surviving relatives. The social insects, including ants, bees, and wasps are most famously studied for this type of relationship because the male drones are clones that share the same genetic make-up as every other male in the colony.[124] In contrast, group selectionists find examples of altruism among non-genetic relatives and explain this through selection acting on the group; whereby, it becomes selectively advantageous for groups if their members express altruistic behaviours to one another. Groups with predominantly altruistic members survive better than groups with predominantly selfish members.[124][136] Coevolution Main article: Coevolution  Bumblebees and the flowers they pollinate have coevolved so that both have become dependent on each other for survival.  Parasitism: A harvestman arachnid being parasitized by mites. The harvestman is being consumed, while the mites benefit from traveling on and feeding off of their host. Ecological interactions can be classified broadly into a host and an associate relationship. A host is any entity that harbours another that is called the associate.[137] Relationships between species that are mutually or reciprocally beneficial are called mutualisms. Examples of mutualism include fungus-growing ants employing agricultural symbiosis, bacteria living in the guts of insects and other organisms, the fig wasp and yucca moth pollination complex, lichens with fungi and photosynthetic algae, and corals with photosynthetic algae.[138][139] If there is a physical connection between host and associate, the relationship is called symbiosis. Approximately 60% of all plants, for example, have a symbiotic relationship with arbuscular mycorrhizal fungi living in their roots forming an exchange network of carbohydrates for mineral nutrients.[140] Indirect mutualisms occur where the organisms live apart. For example, trees living in the equatorial regions of the planet supply oxygen into the atmosphere that sustains species living in distant polar regions of the planet. This relationship is called commensalism because many others receive the benefits of clean air at no cost or harm to trees supplying the oxygen.[5][141] If the associate benefits while the host suffers, the relationship is called parasitism. Although parasites impose a cost to their host (e.g., via damage to their reproductive organs or propagules, denying the services of a beneficial partner), their net effect on host fitness is not necessarily negative and, thus, becomes difficult to forecast.[142][143] Co-evolution is also driven by competition among species or among members of the same species under the banner of reciprocal antagonism, such as grasses competing for growth space. The Red Queen Hypothesis, for example, posits that parasites track down and specialize on the locally common genetic defense systems of its host that drives the evolution of sexual reproduction to diversify the genetic constituency of populations responding to the antagonistic pressure.[144][145] Biogeography Main article: Biogeography Biogeography (an amalgamation of biology and geography) is the comparative study of the geographic distribution of organisms and the corresponding evolution of their traits in space and time.[146] The Journal of Biogeography was established in 1974.[147] Biogeography and ecology share many of their disciplinary roots. For example, the theory of island biogeography, published by the Robert MacArthur and Edward O. Wilson in 1967[148] is considered one of the fundamentals of ecological theory.[149] Biogeography has a long history in the natural sciences concerning the spatial distribution of plants and animals. Ecology and evolution provide the explanatory context for biogeographical studies.[146] Biogeographical patterns result from ecological processes that influence range distributions, such as migration and dispersal.[149] and from historical processes that split populations or species into different areas. The biogeographic processes that result in the natural splitting of species explain much of the modern distribution of the Earth's biota. The splitting of lineages in a species is called vicariance biogeography and it is a sub-discipline of biogeography.[150] There are also practical applications in the field of biogeography concerning ecological systems and processes. For example, the range and distribution of biodiversity and invasive species responding to climate change is a serious concern and active area of research in the context of global warming.[151][152] r/K selection theory Main article: r/K selection theory A population ecology concept is r/K selection theory,[D] one of the first predictive models in ecology used to explain life-history evolution. The premise behind the r/K selection model is that natural selection pressures change according to population density. For example, when an island is first colonized, density of individuals is low. The initial increase in population size is not limited by competition, leaving an abundance of available resources for rapid population growth. These early phases of population growth experience density-independent forces of natural selection, which is called r-selection. As the population becomes more crowded, it approaches the island's carrying capacity, thus forcing individuals to compete more heavily for fewer available resources. Under crowded conditions, the population experiences density-dependent forces of natural selection, called K-selection.[153] In the r/K-selection model, the first variable r is the intrinsic rate of natural increase in population size and the second variable K is the carrying capacity of a population.[33] Different species evolve different life-history strategies spanning a continuum between these two selective forces. An r-selected species is one that has high birth rates, low levels of parental investment, and high rates of mortality before individuals reach maturity. Evolution favours high rates of fecundity in r-selected species. Many kinds of insects and invasive species exhibit r-selected characteristics. In contrast, a K-selected species has low rates of fecundity, high levels of parental investment in the young, and low rates of mortality as individuals mature. Humans and elephants are examples of species exhibiting K-selected characteristics, including longevity and efficiency in the conversion of more resources into fewer offspring.[148][154] Molecular ecology Main article: Molecular ecology The important relationship between ecology and genetic inheritance predates modern techniques for molecular analysis. Molecular ecological research became more feasible with the development of rapid and accessible genetic technologies, such as the polymerase chain reaction (PCR). The rise of molecular technologies and the influx of research questions into this new ecological field resulted in the publication Molecular Ecology in 1992.[155] Molecular ecology uses various analytical techniques to study genes in an evolutionary and ecological context. In 1994, John Avise also played a leading role in this area of science with the publication of his book, Molecular Markers, Natural History and Evolution.[156] Newer technologies opened a wave of genetic analysis into organisms once difficult to study from an ecological or evolutionary standpoint, such as bacteria, fungi, and nematodes. Molecular ecology engendered a new research paradigm for investigating ecological questions considered otherwise intractable. Molecular investigations revealed previously obscured details in the tiny intricacies of nature and improved resolution into probing questions about behavioural and biogeographical ecology.[156] For example, molecular ecology revealed promiscuous sexual behaviour and multiple male partners in tree swallows previously thought to be socially monogamous.[157] In a biogeographical context, the marriage between genetics, ecology, and evolution resulted in a new sub-discipline called phylogeography.[158] |
複雑さ 主な記事 複雑さ も参照のこと: 創発 複雑性とは、相互作用する多数の部分をつなぎ合わせるために必要な、人間の頭の記憶容量を超える大きな計算努力として理解される。生物多様性のグローバル なパターンは複雑である。この生物学的複雑性は、ランドスケープにまたがる過渡的な領域やエコトーンのような、互いに影響し合う異なるスケールでのパター ンに作用する生態学的プロセス間の相互作用から生じている。複雑性は、エネルギーや物質がより大きな単位に統合され、より小さな部分に重なり合うように、 生物学的組織のレベル間の相互作用から生じている。あるレベルでは全体であったものが、より高いレベルでは部分となる」[98]:209 小規模なパターンが必ずしも大規模な現象を説明するわけではなく、そうでなければ(アリストテレスの造語である)「和は部分よりも大きい」という表現にと らえられる[99][100][E]。 「生態学における複雑性には、空間的、時間的、構造的、過程的、行動的、幾何学的という少なくとも6つの異なるタイプがある」[101]: これらの原則 から、生態学者は、分子レベルから惑星レベルに至るまで、影響力の異なる環境スケールで作用する創発的現象や自己組織化現象を同定しており、これらの現象 は、それぞれの統合レベルにおいて異なる説明を必要としている[48][102]。生態系の複雑性は、歴史のランダムな揺らぎによって指示される複数の移 り変わる定常状態に移行する生態系の動的回復力に関連している[9][103]。長期生態学的研究は、より長い時間スケールやより広い空間スケールにおけ る生態系の複雑性と回復力をよりよく理解するための重要な実績を提供する。これらの研究は、国際長期生態学ネットワーク(LTER)によって管理されてい る[104]。現存する最も長い実験は、1856年に開始された公園草実験である[105]。 ホリズム 主な記事 ホリズム ホリズムは、現代の生態学研究における理論的基盤の重要な部分を占めている。ホリズムは、非還元的な特性に従って機能する創発的な全体システムの層へと自 己組織化する生命の生物学的組織を扱っている。これは、生態系のような機能システム全体の高次のパターンは、単純な部分の総和では予測も理解もできないこ とを意味する[107]。「新しい性質が現れるのは、構成要素が相互作用するからであって、構成要素の基本的な性質が変わるからではない」[5]: 8 1)生態系の機械論的な複雑さ、2)相関関係は特定できるかもしれないが、システム全体を参照しなければ因果関係については何も理解できない定量的な還元 主義的用語によるパターンの実際的な記述、3)より小さな部分を参照することなく、より大きなシステムの因果関係を理解する形而上学的な階層。科学的ホリ ズムは、同じ用語を流用した神秘主義とは異なる。形而上学的ホリズムの例は、異なる種の貝殻の外側の厚みが増す傾向に見られる。厚みが増す理由は、外殻の 生体分子特性を参照したり理解したりしなくても、捕食を介した自然淘汰の原理を参照することで理解できる[109]。 +++++++++++++++++++++++++++++++++++++++++++ 進化との関係 主な記事 進化生態学 生態学と進化生物学は生命科学の姉妹分野と考えられている。自然淘汰、生活史、発生、適応、個体群、遺伝などは、生態学と進化論に等しく通じる概念の一例 である。例えば、形態学的、行動学的、遺伝学的形質を進化系統樹にマッピングすることで、異なる生態学的状況における種の機能と役割に関連して、種の歴史 的発展を研究することができる。このような枠組みでは、系統分類学やリンネの分類体系のような共通の体系的原理によって生命を整理、分類、調査するため、 生態学者と進化学者の分析ツールは重なり合う。生態学と進化学の境界は必ずしも明確ではないが、生態学者は進化のプロセスに影響を与える生物学的・生物学 的要因を研究しており[112][113]、進化は1世代という短い生態学的時間スケールで起こることもあり、急速であることもある[114]。 行動生態学 主な記事 行動生態学 カメレオン(Bradypodion spp.)の異なる適応種における社会的ディスプレイと色の変化。カメレオンは行動防御機構として、背景に合わせて皮膚の色を変化させる。また、上の3種 (A-C)に示す支配的(左)対従順(右)のパターンのように、色を使って他の種とコミュニケーションをとることもある[115]。 すべての生物は行動を示すことができる。植物でさえ、記憶やコミュニケーションを含む複雑な行動を示す。行動生態学とは、環境における生物の行動と、その 生態学的・進化的意味を研究する学問である。行動生態学とは、環境における生物の行動と、その生態学的・進化学的意味を研究する学問である。これには、植 物の運動性精子、移動性植物プランクトン、雌の卵に向かって泳ぐ動物プランクトン、ゾウムシによる菌類の培養、サンショウウオの交尾ダンス、アメーバの社 会的集会などの調査が含まれる[117][118][119][120][121]。 適応は行動生態学の中心的な統一概念である。行動も、目や髪の色と同じように、形質として記録され、遺伝することができる。行動は自然淘汰によって、生殖適性を高める機能的有用性を付与する適応形質として進化することができる[123][124]。 相互主義: ヨコバイ(Eurymela fenestrata)はアリ(Iridomyrmex purpureus)によって保護される相互扶助関係にある。アリはヨコバイを捕食者から守り、ヨコバイの摂食を刺激する。その見返りとして、植物を摂食 するヨコバイは肛門からミツロウを分泌し、世話をしているアリにエネルギーと栄養を供給する。 捕食者と被食者の相互作用は、行動生態学だけでなく、食物網の研究においても入門的な概念である。多くの餌生物種は、危険の度合いが異なる複数の捕食者に 直面している。環境に適応し、捕食の脅威に立ち向かうために、生物は成長、摂食、交尾、社会化、生息地の改変など、生活史のさまざまな側面に投資しなが ら、エネルギー収支のバランスをとらなければならない。行動生態学で提唱される仮説は、一般的に保全、最適化、効率化といった適応原則に基づいている。 [33][112][127]例えば、「脅威に敏感な捕食者回避仮説は、獲物は異なる捕食者によってもたらされる脅威の程度を評価し、現在のリスクレベル に応じて行動を一致させるべきであると予測する」[128]、あるいは「最適な飛翔開始距離は、捕食者の初期体力、逃走しないことによって得られる利益、 エネルギー的逃走コスト、捕食リスクによる予想体力損失によって決まる、遭遇後の予想体力が最大になる場所で発生する」[129]。 動物の行動生態学では、精巧な性的ディスプレイやポーズが見られる。例えば、極楽鳥類は求愛の際に歌い、手の込んだ装飾品を見せる。これらのディスプレイ には、健康な個体や適応した個体、望ましい遺伝子を示すという2つの目的がある。このディスプレイは、求婚者間の形質の質の広告として、性淘汰によって駆 動される[130]。 認知生態学 認知生態学は、進化生態学と神経生物学、主に認知科学からの理論と観察を統合し、動物とその生息環境との相互作用が認知システムに与える影響と、それらの システムが生態学的・進化的枠組みの中でどのように行動を制限するかを理解するためのものである。認知に対する淘汰圧を考慮することで、認知生態学は認知 の学際的研究に知的一貫性を貢献することができる」[132][133]。生物と環境の間の「結合」や相互作用を含む研究として、認知生態学は「...我 々は生物と環境を相互仕様と淘汰の中で結合していると見なければならない...」という見解に基づく分野であるenactivism[131]と密接に関 連している[134]。 社会生態学 主な記事 社会生態学(学問分野) 社会生態学的行動は、真社会性が進化した社会性昆虫、粘菌、社会性クモ、人間社会、ハダカデバネズミなどで顕著である。社会行動には、親族間や巣の仲間間 の互恵的な行動が含まれ[119][124][135]、親族選択と集団選択から進化する。血縁淘汰は遺伝的関係を通じて利他主義を説明し、死に至る利他 的行動は、生き残った親族に分配される遺伝的コピーの生存によって報われる。アリ、ハチ、スズメバチなどの社会性昆虫がこの種の関係で最も有名に研究され ているのは、雄のドローンがコロニー内の他のすべての雄と同じ遺伝的構成を共有するクローンであるためである。利他的なメンバーが多い集団は、利己的なメ ンバーが多い集団よりも生存率が高いのである[124][136]。 共進化 主な記事 共進化 マルハナバチと彼らが受粉させる花は共進化し、両者は生存のために互いに依存するようになった。 寄生: ダニに寄生されるハクビシン蜘蛛。ハクビシン蜘蛛は食べられているが、ダニは宿主に寄生して食べることで利益を得ている。 生態学的相互作用は、宿主と仲間の関係に大別できる。宿主とは、仲間(アソシエイト)と呼ばれる別の存在を宿すあらゆる存在のことである。相互作用の例と しては、農業共生を行う菌類生育アリ、昆虫や他の生物の腸内に生息するバクテリア、イチジクコバチとユッカガの受粉複合体、菌類や光合成藻類を持つ地衣 類、光合成藻類を持つサンゴなどが挙げられる[138][139]。例えば、全植物の約60%は、根に生息するアーバスキュラー菌根菌と共生関係にあり、 ミネラル栄養素と炭水化物の交換ネットワークを形成している[140]。 間接的な相互作用は、生物が離れて暮らしている場合に起こる。例えば、地球の赤道直下に生息する樹木は、遠く離れた極地に生息する生物種を維持するため に、大気中に酸素を供給している。酸素を供給している樹木に犠牲や害を与えることなく、他の多くの生物がきれいな空気の恩恵を受けているため、このような 関係は「共生」と呼ばれる。寄生虫は宿主に犠牲を強いるが(例えば、生殖器官や繁殖体にダメージを与えたり、有益なパートナーのサービスを拒否したりする ことによって)、宿主のフィットネスに対する正味の影響は必ずしもマイナスではなく、したがって予測は困難となる。例えば、赤の女王仮説は、寄生虫が宿主 の局所的に共通な遺伝的防御システムを突き止め、それに特化することで、有性生殖の進化が促進され、拮抗的な圧力に対応する個体群の遺伝的構成要素が多様 化すると仮定している[144][145]。 生物地理学 主な記事 生物地理学 生物地理学(生物学と地理学の融合)とは、生物の地理的分布とそれに対応する形質の進化を空間的・時間的に比較研究する学問である[146]。生物地理学 雑誌は1974年に創刊された[147]。例えば、1967年にロバート・マッカーサーとエドワード・O・ウィルソンによって発表された島嶼生物地理学の 理論[148]は、生態学理論の基礎の一つと考えられている[149]。 生物地理学は、動植物の空間分布に関する自然科学において長い歴史を持っている。生物地理学的パターンは、移動や分散といった範囲分布に影響を与える生態 学的プロセス[149]や、個体群や種を異なる地域に分割する歴史的プロセスから生じる。種の自然分裂をもたらす生物地理学的プロセスは、地球の生物相の 現代的分布の多くを説明している。種における系統の分裂は、迂回生物地理学と呼ばれ、生物地理学の下位分野のひとつである。例えば、気候変動に対応する生 物多様性や侵入種の範囲や分布は、地球温暖化の文脈における重大な関心事であり、活発な研究分野である[151][152]。 r/K選択理論 主な記事:r/K選択理論 集団生態学の概念にr/K選択理論[D]があり、生命誌進化を説明するために使われた生態学における最初の予測モデルの一つである。r/K選択モデルの前 提となっているのは、自然選択圧は個体群密度によって変化するということである。例えば、ある島が最初に植民地化されたとき、個体密度は低い。初期の個体 数の増加は競争によって制限されることはなく、利用可能な資源は豊富にあるため、個体数は急速に増加する。このような人口増加の初期段階には、密度に依存 しない自然淘汰の力が働き、これはr-淘汰と呼ばれる。個体群が混雑してくると、島の収容力に近づくため、個体はより少ない利用可能な資源をより激しく奪 い合うことになる。混雑した条件下では、集団はK-選択と呼ばれる密度依存的な自然選択の力を経験する[153]。 r/K-選択モデルでは、最初の変数rは個体数の自然増加率であり、2番目の変数Kは個体群の環境収容力である。r選択種とは、出生率が高く、親への投資 レベルが低く、個体が成熟するまでの死亡率が高い種である。進化はr-選択種の高い繁殖率を好む。多くの昆虫や外来種がr-選択性の特徴を示している。こ れとは対照的に、K-選択種は繁殖率が低く、子供への親からの投資が多く、個体が成熟するにつれて死亡率が低くなる。ヒトやゾウは、長寿や、より多くの資 源をより少ない子孫に変換する効率性など、K-選択的特徴を示す種の例である[148][154]。 分子生態学 主な記事 分子生態学 生態学と遺伝の間の重要な関係は、分子解析のための近代的な技術以前からあった。分子生態学的研究は、ポリメラーゼ連鎖反応(PCR)のような、迅速でア クセスしやすい遺伝学的技術の発展とともに、より実現可能なものとなった。分子技術の台頭とこの新しい生態学分野への研究課題の流入により、1992年に 『分子生態学』が出版された[155]。分子生態学は、遺伝子を進化的・生態学的な文脈で研究するために様々な分析技術を用いる。1994年、ジョン・ア ヴィーズもまた、著書『Molecular Markers, Natural History and Evolution(分子マーカー、自然史と進化)』を出版し、この分野の科学において主導的な役割を果たした[156]。新しい技術は、バクテリア、真 菌、線虫など、かつては生態学的・進化学的見地から研究することが困難であった生物に対する遺伝子解析の波を開いた。分子生態学は、これまで困難であった と考えられていた生態学的問題を調査するための新しい研究パラダイムを生み出した。例えば、分子生態学は、以前は社会的に一夫一婦制であると考えられてい たツバメの乱交性行動と複数の雄のパートナーを明らかにした[157]。 |
| Human ecology Main article: Human ecology The history of life on Earth has been a history of interaction between living things and their surroundings. To a large extent, the physical form and the habits of the earth's vegetation and its animal life have been molded by the environment. Considering the whole span of earthly time, the opposite effect, in which life actually modifies its surroundings, has been relatively slight. Only within the moment of time represented by the present century has one species man acquired significant power to alter the nature of his world. Rachel Carson, "Silent Spring"[159] Ecology is as much a biological science as it is a human science.[5] Human ecology is an interdisciplinary investigation into the ecology of our species. "Human ecology may be defined: (1) from a bioecological standpoint as the study of man as the ecological dominant in plant and animal communities and systems; (2) from a bioecological standpoint as simply another animal affecting and being affected by his physical environment; and (3) as a human being, somehow different from animal life in general, interacting with physical and modified environments in a distinctive and creative way. A truly interdisciplinary human ecology will most likely address itself to all three."[160]: 3 The term was formally introduced in 1921, but many sociologists, geographers, psychologists, and other disciplines were interested in human relations to natural systems centuries prior, especially in the late 19th century.[160][161] The ecological complexities human beings are facing through the technological transformation of the planetary biome has brought on the Anthropocene. The unique set of circumstances has generated the need for a new unifying science called coupled human and natural systems that builds upon, but moves beyond the field of human ecology.[107] Ecosystems tie into human societies through the critical and all-encompassing life-supporting functions they sustain. In recognition of these functions and the incapability of traditional economic valuation methods to see the value in ecosystems, there has been a surge of interest in social-natural capital, which provides the means to put a value on the stock and use of information and materials stemming from ecosystem goods and services. Ecosystems produce, regulate, maintain, and supply services of critical necessity and beneficial to human health (cognitive and physiological), economies, and they even provide an information or reference function as a living library giving opportunities for science and cognitive development in children engaged in the complexity of the natural world. Ecosystems relate importantly to human ecology as they are the ultimate base foundation of global economics as every commodity, and the capacity for exchange ultimately stems from the ecosystems on Earth.[107][162][163][164] Main article: Restoration ecology See also: Natural resource management Ecosystem management is not just about science nor is it simply an extension of traditional resource management; it offers a fundamental reframing of how humans may work with nature. Grumbine (1994)[165]: 27 Ecology is an employed science of restoration, repairing disturbed sites through human intervention, in natural resource management, and in environmental impact assessments. Edward O. Wilson predicted in 1992 that the 21st century "will be the era of restoration in ecology".[166] Ecological science has boomed in the industrial investment of restoring ecosystems and their processes in abandoned sites after disturbance. Natural resource managers, in forestry, for example, employ ecologists to develop, adapt, and implement ecosystem based methods into the planning, operation, and restoration phases of land-use. Another example of conservation is seen on the east coast of the United States in Boston, MA. The city of Boston implemented the Wetland Ordinance,[167] improving the stability of their wetland environments by implementing soil amendments that will improve groundwater storage and flow, and trimming or removal of vegetation that could cause harm to water quality.[citation needed] Ecological science is used in the methods of sustainable harvesting, disease, and fire outbreak management, in fisheries stock management, for integrating land-use with protected areas and communities, and conservation in complex geo-political landscapes.[22][165][168][169] |
ヒューマンエコロジー 主な記事 人間生態学 地球上の生命の歴史は、生物とその周囲の環境との相互作用の歴史であった。地球の植生や動物たちの物理的な形態や習性は、その大部分において環境によって 形成されてきた。地球上の時間全体を考えると、生命が周囲の環境を実際に変化させるという逆の効果は、比較的わずかなものであった。今世紀に代表される一 瞬の時間の中でだけ、ある種の人間が世界の自然を変える大きな力を手に入れたのである。 レイチェル・カーソン『沈黙の春』[159]。 生態学は生物学であると同時に人間科学でもある。「(2)生物生態学的見地からは、単に物理的環境に影響を与え、またその影響を受けているもう一つの動物 として、(3)一般的な動物とはどこか異なり、独特かつ創造的な方法で物理的環境および改変された環境と相互作用している人間として、である。真に学際的 なヒューマンエコロジーは、おそらくこの3つすべてに取り組むことになるだろう」[160]: 3 この用語は1921年に正式に導入されたが、多くの社会学者、地理学者、心理学者、その他の学問分野は、数世紀前、特に19世紀後半から、自然と人間の関 係に関心を持っていた[160][161]。 惑星生物圏の技術的変容を通じて人類が直面している生態学的複雑性は、人新世をもたらした。生態系は、それらが維持する重要かつ包括的な生命維持機能を通 じて、人間社会と結びついている。このような機能と、従来の経済評価手法では生態系に価値を見出すことができないことを認識し、生態系の財やサービスに由 来する情報や物質のストックや利用に価値を置く手段を提供する社会自然資本への関心が急増している。生態系は、人間の健康(認知的・生理的)、経済にとっ て極めて必要で有益なサービスを生産、調整、維持、供給し、さらには、複雑な自然界に関わる子供たちに科学と認知の発達の機会を与える生きた図書館とし て、情報や参考の機能を提供している。生態系は、あらゆる商品と交換能力が最終的に地球上の生態系に由来するように、グローバル経済の究極的な基盤である ため、人間の生態系と重要な関係がある[107][162][163][164]。 主な記事 修復生態学 以下も参照のこと: 天然資源管理 生態系管理は単なる科学ではなく、伝統的な資源管理の延長でもない。 グランバイン(1994)[165]: 27 生態学は、自然資源管理や環境影響評価において、人間の介入によって乱された場所を修復する、修復の科学として採用されている。エドワード・O・ウィルソ ンは1992年に、21世紀は「生態学における修復の時代」になるだろうと予言している[166]。生態学は、攪乱後に放棄された場所の生態系とそのプロ セスを修復するという産業投資においてブームとなっている。例えば林業などの天然資源管理者は、土地利用の計画、運営、修復の各段階において、生態系に基 づく手法を開発し、適応させ、実施するために生態学者を雇用している。保全のもう一つの例は、アメリカ東海岸のマサチューセッツ州ボストンで見られる。ボ ストン市は湿地条例を施行し[167]、地下水の貯留と流れを改善する土壌改良を実施したり、水質に害を及ぼす可能性のある植生を刈り込んだり除去したり することで、湿地環境の安定性を向上させている[要出典]。生態科学は、持続可能な収穫、疾病、火災の発生管理、漁業資源管理、保護地域やコミュニティと 土地利用の統合、複雑な地政学的ランドスケープにおける保全の手法に用いられている[22][165][168][169]。 |
| Relation to the environment Main article: Natural environment The environment of ecosystems includes both physical parameters and biotic attributes. It is dynamically interlinked and contains resources for organisms at any time throughout their life cycle.[5][170] Like ecology, the term environment has different conceptual meanings and overlaps with the concept of nature. Environment "includes the physical world, the social world of human relations and the built world of human creation."[171]: 62 The physical environment is external to the level of biological organization under investigation, including abiotic factors such as temperature, radiation, light, chemistry, climate and geology. The biotic environment includes genes, cells, organisms, members of the same species (conspecifics) and other species that share a habitat.[172] The distinction between external and internal environments, however, is an abstraction parsing life and environment into units or facts that are inseparable in reality. There is an interpenetration of cause and effect between the environment and life. The laws of thermodynamics, for example, apply to ecology by means of its physical state. With an understanding of metabolic and thermodynamic principles, a complete accounting of energy and material flow can be traced through an ecosystem. In this way, the environmental and ecological relations are studied through reference to conceptually manageable and isolated material parts. After the effective environmental components are understood through reference to their causes; however, they conceptually link back together as an integrated whole, or holocoenotic system as it was once called. This is known as the dialectical approach to ecology. The dialectical approach examines the parts but integrates the organism and the environment into a dynamic whole (or umwelt). Change in one ecological or environmental factor can concurrently affect the dynamic state of an entire ecosystem.[36][173] Disturbance and resilience Main article: Resilience (ecology) Ecosystems are regularly confronted with natural environmental variations and disturbances over time and geographic space. A disturbance is any process that removes biomass from a community, such as a fire, flood, drought, or predation.[174] Disturbances occur over vastly different ranges in terms of magnitudes as well as distances and time periods,[175] and are both the cause and product of natural fluctuations in death rates, species assemblages, and biomass densities within an ecological community. These disturbances create places of renewal where new directions emerge from the patchwork of natural experimentation and opportunity.[174][176][177] Ecological resilience is a cornerstone theory in ecosystem management. Biodiversity fuels the resilience of ecosystems acting as a kind of regenerative insurance.[177] Metabolism and the early atmosphere Main article: Early atmosphere Metabolism – the rate at which energy and material resources are taken up from the environment, transformed within an organism, and allocated to maintenance, growth and reproduction – is a fundamental physiological trait. Ernest et al.[178]: 991 The Earth was formed approximately 4.5 billion years ago.[179] As it cooled and a crust and oceans formed, its atmosphere transformed from being dominated by hydrogen to one composed mostly of methane and ammonia. Over the next billion years, the metabolic activity of life transformed the atmosphere into a mixture of carbon dioxide, nitrogen, and water vapor. These gases changed the way that light from the sun hit the Earth's surface and greenhouse effects trapped heat. There were untapped sources of free energy within the mixture of reducing and oxidizing gasses that set the stage for primitive ecosystems to evolve and, in turn, the atmosphere also evolved.[180] The leaf is the primary site of photosynthesis in many higher plants. Throughout history, the Earth's atmosphere and biogeochemical cycles have been in a dynamic equilibrium with planetary ecosystems. The history is characterized by periods of significant transformation followed by millions of years of stability.[181] The evolution of the earliest organisms, likely anaerobic methanogen microbes, started the process by converting atmospheric hydrogen into methane (4H2 + CO2 → CH4 + 2H2O). Anoxygenic photosynthesis reduced hydrogen concentrations and increased atmospheric methane, by converting hydrogen sulfide into water or other sulfur compounds (for example, 2H2S + CO2 + hv → CH2O + H2O + 2S). Early forms of fermentation also increased levels of atmospheric methane. The transition to an oxygen-dominant atmosphere (the Great Oxidation) did not begin until approximately 2.4–2.3 billion years ago, but photosynthetic processes started 0.3 to 1 billion years prior.[181][182] Radiation: heat, temperature and light The biology of life operates within a certain range of temperatures. Heat is a form of energy that regulates temperature. Heat affects growth rates, activity, behaviour, and primary production. Temperature is largely dependent on the incidence of solar radiation. The latitudinal and longitudinal spatial variation of temperature greatly affects climates and consequently the distribution of biodiversity and levels of primary production in different ecosystems or biomes across the planet. Heat and temperature relate importantly to metabolic activity. Poikilotherms, for example, have a body temperature that is largely regulated and dependent on the temperature of the external environment. In contrast, homeotherms regulate their internal body temperature by expending metabolic energy.[112][113][173] There is a relationship between light, primary production, and ecological energy budgets. Sunlight is the primary input of energy into the planet's ecosystems. Light is composed of electromagnetic energy of different wavelengths. Radiant energy from the sun generates heat, provides photons of light measured as active energy in the chemical reactions of life, and also acts as a catalyst for genetic mutation.[112][113][173] Plants, algae, and some bacteria absorb light and assimilate the energy through photosynthesis. Organisms capable of assimilating energy by photosynthesis or through inorganic fixation of H2S are autotrophs. Autotrophs—responsible for primary production—assimilate light energy which becomes metabolically stored as potential energy in the form of biochemical enthalpic bonds.[112][113][173] Physical environments Water Main article: Aquatic ecosystem Wetland conditions such as shallow water, high plant productivity, and anaerobic substrates provide a suitable environment for important physical, biological, and chemical processes. Because of these processes, wetlands play a vital role in global nutrient and element cycles. Cronk & Fennessy (2001)[183]: 29 Diffusion of carbon dioxide and oxygen is approximately 10,000 times slower in water than in air. When soils are flooded, they quickly lose oxygen, becoming hypoxic (an environment with O2 concentration below 2 mg/liter) and eventually completely anoxic where anaerobic bacteria thrive among the roots. Water also influences the intensity and spectral composition of light as it reflects off the water surface and submerged particles.[183] Aquatic plants exhibit a wide variety of morphological and physiological adaptations that allow them to survive, compete, and diversify in these environments. For example, their roots and stems contain large air spaces (aerenchyma) that regulate the efficient transportation of gases (for example, CO2 and O2) used in respiration and photosynthesis. Salt water plants (halophytes) have additional specialized adaptations, such as the development of special organs for shedding salt and osmoregulating their internal salt (NaCl) concentrations, to live in estuarine, brackish, or oceanic environments. Anaerobic soil microorganisms in aquatic environments use nitrate, manganese ions, ferric ions, sulfate, carbon dioxide, and some organic compounds; other microorganisms are facultative anaerobes and use oxygen during respiration when the soil becomes drier. The activity of soil microorganisms and the chemistry of the water reduces the oxidation-reduction potentials of the water. Carbon dioxide, for example, is reduced to methane (CH4) by methanogenic bacteria.[183] The physiology of fish is also specially adapted to compensate for environmental salt levels through osmoregulation. Their gills form electrochemical gradients that mediate salt excretion in salt water and uptake in fresh water.[184] Gravity The shape and energy of the land are significantly affected by gravitational forces. On a large scale, the distribution of gravitational forces on the earth is uneven and influences the shape and movement of tectonic plates as well as influencing geomorphic processes such as orogeny and erosion. These forces govern many of the geophysical properties and distributions of ecological biomes across the Earth. On the organismal scale, gravitational forces provide directional cues for plant and fungal growth (gravitropism), orientation cues for animal migrations, and influence the biomechanics and size of animals.[112] Ecological traits, such as allocation of biomass in trees during growth are subject to mechanical failure as gravitational forces influence the position and structure of branches and leaves.[185] The cardiovascular systems of animals are functionally adapted to overcome the pressure and gravitational forces that change according to the features of organisms (e.g., height, size, shape), their behaviour (e.g., diving, running, flying), and the habitat occupied (e.g., water, hot deserts, cold tundra).[186] Pressure Climatic and osmotic pressure places physiological constraints on organisms, especially those that fly and respire at high altitudes, or dive to deep ocean depths.[187] These constraints influence vertical limits of ecosystems in the biosphere, as organisms are physiologically sensitive and adapted to atmospheric and osmotic water pressure differences.[112] For example, oxygen levels decrease with decreasing pressure and are a limiting factor for life at higher altitudes.[188] Water transportation by plants is another important ecophysiological process affected by osmotic pressure gradients.[189][190][191] Water pressure in the depths of oceans requires that organisms adapt to these conditions. For example, diving animals such as whales, dolphins, and seals are specially adapted to deal with changes in sound due to water pressure differences.[192] Differences between hagfish species provide another example of adaptation to deep-sea pressure through specialized protein adaptations.[193] Wind and turbulence The architecture of the inflorescence in grasses is subject to the physical pressures of wind and shaped by the forces of natural selection facilitating wind-pollination (anemophily).[194][195] Turbulent forces in air and water affect the environment and ecosystem distribution, form, and dynamics. On a planetary scale, ecosystems are affected by circulation patterns in the global trade winds. Wind power and the turbulent forces it creates can influence heat, nutrient, and biochemical profiles of ecosystems.[112] For example, wind running over the surface of a lake creates turbulence, mixing the water column and influencing the environmental profile to create thermally layered zones, affecting how fish, algae, and other parts of the aquatic ecosystem are structured.[196][197] Wind speed and turbulence also influence evapotranspiration rates and energy budgets in plants and animals.[183][198] Wind speed, temperature and moisture content can vary as winds travel across different land features and elevations. For example, the westerlies come into contact with the coastal and interior mountains of western North America to produce a rain shadow on the leeward side of the mountain. The air expands and moisture condenses as the winds increase in elevation; this is called orographic lift and can cause precipitation. This environmental process produces spatial divisions in biodiversity, as species adapted to wetter conditions are range-restricted to the coastal mountain valleys and unable to migrate across the xeric ecosystems (e.g., of the Columbia Basin in western North America) to intermix with sister lineages that are segregated to the interior mountain systems.[199][200] Fire Main article: Fire ecology Forest fires modify the land by leaving behind an environmental mosaic that diversifies the landscape into different seral stages and habitats of varied quality (left). Some species are adapted to forest fires, such as pine trees that open their cones only after fire exposure (right). Plants convert carbon dioxide into biomass and emit oxygen into the atmosphere. By approximately 350 million years ago (the end of the Devonian period), photosynthesis had brought the concentration of atmospheric oxygen above 17%, which allowed combustion to occur.[201] Fire releases CO2 and converts fuel into ash and tar. Fire is a significant ecological parameter that raises many issues pertaining to its control and suppression.[202] While the issue of fire in relation to ecology and plants has been recognized for a long time,[203] Charles Cooper brought attention to the issue of forest fires in relation to the ecology of forest fire suppression and management in the 1960s.[204][205] Native North Americans were among the first to influence fire regimes by controlling their spread near their homes or by lighting fires to stimulate the production of herbaceous foods and basketry materials.[206] Fire creates a heterogeneous ecosystem age and canopy structure, and the altered soil nutrient supply and cleared canopy structure opens new ecological niches for seedling establishment.[207][208] Most ecosystems are adapted to natural fire cycles. Plants, for example, are equipped with a variety of adaptations to deal with forest fires. Some species (e.g., Pinus halepensis) cannot germinate until after their seeds have lived through a fire or been exposed to certain compounds from smoke. Environmentally triggered germination of seeds is called serotiny.[209][210] Fire plays a major role in the persistence and resilience of ecosystems.[176] Soils Main article: Soil ecology Soil is the living top layer of mineral and organic dirt that covers the surface of the planet. It is the chief organizing centre of most ecosystem functions, and it is of critical importance in agricultural science and ecology. The decomposition of dead organic matter (for example, leaves on the forest floor), results in soils containing minerals and nutrients that feed into plant production. The whole of the planet's soil ecosystems is called the pedosphere where a large biomass of the Earth's biodiversity organizes into trophic levels. Invertebrates that feed and shred larger leaves, for example, create smaller bits for smaller organisms in the feeding chain. Collectively, these organisms are the detritivores that regulate soil formation.[211][212] Tree roots, fungi, bacteria, worms, ants, beetles, centipedes, spiders, mammals, birds, reptiles, amphibians, and other less familiar creatures all work to create the trophic web of life in soil ecosystems. Soils form composite phenotypes where inorganic matter is enveloped into the physiology of a whole community. As organisms feed and migrate through soils they physically displace materials, an ecological process called bioturbation. This aerates soils and stimulates heterotrophic growth and production. Soil microorganisms are influenced by and are fed back into the trophic dynamics of the ecosystem. No single axis of causality can be discerned to segregate the biological from geomorphological systems in soils.[213][214] Paleoecological studies of soils places the origin for bioturbation to a time before the Cambrian period. Other events, such as the evolution of trees and the colonization of land in the Devonian period played a significant role in the early development of ecological trophism in soils.[212][215][216] Biogeochemistry and climate Main article: Biogeochemistry See also: Nutrient cycle and Climate Ecologists study and measure nutrient budgets to understand how these materials are regulated, flow, and recycled through the environment.[112][113][173] This research has led to an understanding that there is global feedback between ecosystems and the physical parameters of this planet, including minerals, soil, pH, ions, water, and atmospheric gases. Six major elements (hydrogen, carbon, nitrogen, oxygen, sulfur, and phosphorus; H, C, N, O, S, and P) form the constitution of all biological macromolecules and feed into the Earth's geochemical processes. From the smallest scale of biology, the combined effect of billions upon billions of ecological processes amplify and ultimately regulate the biogeochemical cycles of the Earth. Understanding the relations and cycles mediated between these elements and their ecological pathways has significant bearing toward understanding global biogeochemistry.[217] The ecology of global carbon budgets gives one example of the linkage between biodiversity and biogeochemistry. It is estimated that the Earth's oceans hold 40,000 gigatonnes (Gt) of carbon, that vegetation and soil hold 2070 Gt, and that fossil fuel emissions are 6.3 Gt carbon per year.[218] There have been major restructurings in these global carbon budgets during the Earth's history, regulated to a large extent by the ecology of the land. For example, through the early-mid Eocene volcanic outgassing, the oxidation of methane stored in wetlands, and seafloor gases increased atmospheric CO2 (carbon dioxide) concentrations to levels as high as 3500 ppm.[219] In the Oligocene, from twenty-five to thirty-two million years ago, there was another significant restructuring of the global carbon cycle as grasses evolved a new mechanism of photosynthesis, C4 photosynthesis, and expanded their ranges. This new pathway evolved in response to the drop in atmospheric CO2 concentrations below 550 ppm.[220] The relative abundance and distribution of biodiversity alters the dynamics between organisms and their environment such that ecosystems can be both cause and effect in relation to climate change. Human-driven modifications to the planet's ecosystems (e.g., disturbance, biodiversity loss, agriculture) contributes to rising atmospheric greenhouse gas levels. Transformation of the global carbon cycle in the next century is projected to raise planetary temperatures, lead to more extreme fluctuations in weather, alter species distributions, and increase extinction rates. The effect of global warming is already being registered in melting glaciers, melting mountain ice caps, and rising sea levels. Consequently, species distributions are changing along waterfronts and in continental areas where migration patterns and breeding grounds are tracking the prevailing shifts in climate. Large sections of permafrost are also melting to create a new mosaic of flooded areas having increased rates of soil decomposition activity that raises methane (CH4) emissions. There is concern over increases in atmospheric methane in the context of the global carbon cycle, because methane is a greenhouse gas that is 23 times more effective at absorbing long-wave radiation than CO2 on a 100-year time scale.[221] Hence, there is a relationship between global warming, decomposition and respiration in soils and wetlands producing significant climate feedbacks and globally altered biogeochemical cycles.[107][222][223][224][225][226] |
環境との関係 主な記事 自然環境 生態系の環境には、物理的パラメータと生物学的属性の両方が含まれる。生態学と同様に、環境という用語は異なる概念的意味を持ち、自然という概念と重複し ている。環境は「物理的世界、人間関係の社会的世界、人間が創造した建造物の世界を含む」[171]。物理的環境は、温度、放射線、光、化学、気候、地質 などの生物学的要因を含む、調査対象の生物学的組織レベルの外部にある。生物学的環境には、遺伝子、細胞、生物、同じ種のメンバー(同種)、生息地を共有 する他の種が含まれる[172]。 しかし、外部環境と内部環境の区別は、生命と環境を現実には切り離せない単位や事実に分節化した抽象的なものである。環境と生命の間には、原因と結果の相 互浸透がある。例えば、熱力学の法則は、その物理的状態によって生態系に適用される。代謝と熱力学の原理を理解すれば、生態系を通してエネルギーと物質の 流れを完全に把握することができる。このようにして、環境と生態系の関係は、概念的に管理可能で分離された物質部分を参照することによって研究される。効 果的な環境構成要素は、その原因を参照することで理解される。しかし、それらは概念的に統合された全体として、あるいはかつてホロコエノティック・システ ムと呼ばれたように、再び結びつけられる。これは生態学の弁証法的アプローチとして知られている。弁証法的アプローチでは、各部分を検証するが、生物と環 境をダイナミックな全体(またはウムヴェルト)として統合する。1つの生態学的要因や環境要因の変化は、同時に生態系全体の動的状態に影響を及ぼす可能性 がある。 撹乱と回復力 主な記事 レジリエンス(生態学) 生態系は、時間的・地理的空間的な自然の環境変動や撹乱に定期的に直面している。攪乱とは、火災、洪水、干ばつ、捕食など、生物群集からバイオマスを除去 するあらゆるプロセスのことである[174]。攪乱は、距離や期間だけでなく、大きさの点でも大きく異なる範囲にわたって発生し[175]、生態系群集内 の死亡率、種群、バイオマス密度における自然変動の原因であると同時に産物でもある。こうした攪乱は、自然の実験と機会のパッチワークから新たな方向性が 生まれる再生の場を生み出す[174][176][177]。生態系の回復力は、生態系管理における基礎理論である。生物多様性は、一種の再生保険として 機能する生態系の回復力を促進する[177]。 代謝と初期大気 主な記事 初期大気 代謝(エネルギーと物質資源が環境から取り込まれ、生物内で変換され、維持、成長、生殖に割り当てられる速度)は、基本的な生理学的特性である。 アーネスト他[178]: 991 地球が冷え、地殻と海洋が形成されるにつれて、その大気は水素を主成分とするものから、メタンとアンモニアを主成分とするものへと変化した。その後10億 年の間に、生命の代謝活動によって、大気は二酸化炭素、窒素、水蒸気の混合物に変化した。これらのガスは、太陽からの光が地表に当たる方法を変え、温室効 果によって熱を閉じ込めた。還元性ガスと酸化性ガスの混合物の中には、原始的な生態系が進化するための舞台を整え、ひいては大気も進化するための、未開発 の自由エネルギー源があった[180]。 葉は、多くの高等植物における光合成の主要な場所である。 歴史を通じて、地球の大気と生物地球化学サイクルは、惑星の生態系と動的な均衡を保ってきた。その歴史は、数百万年の安定に続く、大きな変革の時期によっ て特徴づけられる[181]。おそらく嫌気性メタン生成微生物と思われる、最も初期の生物の進化は、大気中の水素をメタンに変換することによってプロセス を開始した(4H2 + CO2 → CH4 + 2H2O)。無酸素性光合成は、硫化水素を水や他の硫黄化合物に変換することで、水素濃度を低下させ、大気中のメタンを増加させた(例えば、2H2S + CO2 + hv → CH2O + H2O + 2S)。初期の発酵形態も、大気中のメタン濃度を増加させた。酸素優勢大気への移行(大酸化)は約24億~23億年前まで始まらなかったが、光合成プロセ スはその3億~10億年前に始まっていた[181][182]。 放射線:熱、温度、光 生命の生物学は、一定の温度範囲内で活動する。熱は温度を調節するエネルギーの一形態である。熱は成長率、活動、行動、一次生産に影響を与える。温度は太 陽放射の入射に大きく左右される。気温の緯度と経度の空間的変動は気候に大きく影響し、その結果、地球上のさまざまな生態系やバイオームにおける生物多様 性の分布や一次生産のレベルに影響を与える。熱と温度は代謝活動に大きく関係している。例えば、恒温動物は体温を調節し、外部環境の温度に大きく依存す る。一方、恒温動物は代謝エネルギーを消費することで体内温度を調節している[112][113][173]。 光、一次生産、生態学的エネルギー収支の間には関係がある。太陽光は、地球の生態系へのエネルギーの主要な投入源である。光は、異なる波長の電磁エネル ギーで構成されている。太陽からの放射エネルギーは熱を発生させ、生命の化学反応における活性エネルギーとして測定される光の光子を提供し、また遺伝子の 突然変異の触媒として働く[112][113][173]。光合成またはH2Sの無機固定によってエネルギーを同化できる生物は、独立栄養生物である。一 次生産に責任を持つ独立栄養生物は、光エネルギーを同化し、その光エネルギーは、生化学的エンタルピー結合の形で位置エネルギーとして代謝的に蓄積される [112][113][173]。 物理的環境 水 主な記事 水生生態系 浅瀬、高い植物生産性、および嫌気性基質などの湿地の条件は、重要な物理的、生物学的、および化学的プロセスに適した環境を提供する。これらのプロセスにより、湿地は、世界の栄養塩類と元素の循環において重要な役割を果た している。 Cronk & Fennessy (2001)[183]: 29 二酸化炭素と酸素の拡散は、水中では空気中よりも約 10,000 倍遅い。土壌が湛水されると、土壌は急速に酸素を失い、低酸素状態(酸素濃度が 2 mg/l 未満の環境)になり、最終的には完全に無酸素状態になり、嫌気性細菌が根の中で繁殖する。水はまた、水面や水中粒子に反射する光の強度やスペクトル組成に も影響を与える[183]。水生植物は、このような環境で生き残り、競争し、多様化するために、様々な形態学的・生理学的適応を示す。例えば、根や茎には 大きな空隙(aerenchyma)があり、呼吸や光合成に使用される気体(CO2やO2など)の効率的な輸送を制御している。海水植物(塩生植物)は、 河口域、汽水域、海洋環境で生息するために、塩分を排出するための特別な器官を発達させたり、体内の塩分(NaCl)濃度を調節するなど、さらに特殊な適 応を持っている。水生環境の嫌気性土壌微生物は、硝酸塩、マンガンイオン、鉄イオン、硫酸塩、二酸化炭素、およびいくつかの有機化合物を利用する。他の微 生物は通性嫌気性であり、土壌が乾燥すると呼吸の際に酸素を利用する。土壌微生物の活動と水の化学的性質は、水の酸化還元電位を低下させる。例えば二酸化 炭素は、メタン生成細菌によってメタン(CH4)に還元される[183]。魚類の生理学もまた、浸透調節によって環境の塩分濃度を補正するために特別に適 応している。魚の鰓は電気化学的勾配を形成し、海水中では塩分を排泄し、淡水中では塩分を取り込む。 重力 陸地の形状とエネルギーは、重力の影響を大きく受ける。大規模なスケールでは、地球上の重力の分布は不均一であり、造山や侵食などの地形形成過程に影響を 及ぼすだけでなく、地殻プレートの形や動きにも影響を及ぼす。これらの力は、地球全体の多くの地球物理学的特性と生態系バイオームの分布を支配している。 生物スケールでは、重力は植物や菌類の成長(重力屈性)、動物の移動のための方位の手がかりを提供し、動物のバイオメカニクスやサイズに影響を与える [112]。重力が枝や葉の位置や構造に影響を与えるため、成長中の樹木におけるバイオマスの配分などの生態学的形質は、機械的故障の対象となる [185]、 動物の心臓血管系は、生物の特徴(身長、大きさ、形状など)、行動(潜水、走行、飛行など)、生息環境(水、高温の砂漠、低温のツンドラなど)に応じて変 化する圧力と重力に打ち勝つために機能的に適応している。] 圧力 気候圧力と浸透圧は、生物、特に高高度を飛行し呼吸する生物、または深海深度まで潜水する生物に生理的制約を与える。生物は大気圧と浸透圧の水圧差に生理 的に敏感であり適応しているため、これらの制約は生物圏における生態系の垂直限界に影響を及ぼす。 [例えば、酸素濃度は気圧の低下とともに減少し、高高度の生物にとって制限因子となる[188]。植物による水輸送も、浸透圧勾配の影響を受ける重要な生 態生理学的プロセスである[189][190][191]。深海における水圧は、生物にこれらの条件への適応を要求する。例えば、クジラ、イルカ、アザラ シなどの潜水動物は、水圧差による音の変化に対処するために特別に適応している[192]。タガメの種間の違いは、特殊なタンパク質の適応による深海の水 圧への適応のもう一つの例を示している[193]。 風と乱流 イネ科植物の花序の構造は風の物理的圧力を受け、風媒受粉(風媒受粉)を促進する自然選択の力によって形成される[194][195]。 大気と水の乱流力は、環境と生態系の分布、形態、および動態に影響を与える。惑星規模では、生態系は地球規模の貿易風の循環パターンの影響を受ける。風力 とそれが生み出す乱流力は、生態系の熱、栄養塩類、および生化学的プロフィールに影響を与える可能性がある[112]。例えば、湖の水面を流れる風は乱流 を生み出し、水柱を混合し、環境プロフィールに影響を与えて熱層ゾーンを形成し、魚類、藻類、および水生生態系の他の部分の構造に影響を与える。 [196][197]風速と乱流はまた、動植物の蒸発散速度とエネル ギー収支にも影響を与える[183][198]。風速、温度、および含水率は、 風が異なる地形や標高を通過する際に変化する可能性がある。例えば、偏西風は北アメリカ西部の沿岸部や内陸部の山々に接触し、山の風下側に雨影を作り出 す。これはオログラフィック・リフトと呼ばれ、降水の原因となる。この環境プロセスは、生物多様性に空間的な分断をもたらす。湿潤な条件に適応した種は、 沿岸の山岳渓谷に生息域を限定され、内陸の山岳系に隔離された姉妹系統と交雑するために、乾燥した生態系(例えば、北米西部のコロンビア盆地)を移動する ことができないからである[199][200]。 火災 主な記事 火災生態学 森林火災は、景観を様々な鋸歯状期と様々な質の生息地に多様化させる環境モザイクを残すことで土地を改変する(左)。火災にさらされた後にのみ球果を開くマツのように、森林火災に適応する種もある(右)。 植物は二酸化炭素をバイオマスに変換し、大気中に酸素を放出する。約3億5,000万年前(デボン紀の終わり)には、光合成によって大気中の酸素濃度が 17%以上になり、燃焼が可能になった[201]。火は二酸化炭素を放出し、燃料を灰とタールに変える。火は重要な生態学的パラメータであり、その制御と 抑制に関する多くの問題を提起している[202]。生態学と植物との関係における火の問題は長い間認識されてきたが[203]、チャールズ・クーパーは 1960年代に森林火災の抑制と管理の生態学との関係において森林火災の問題に注目させた[204][205]。 北米先住民は、自宅付近での延焼を制御したり、草食動物や籠の材料の生産を刺激するために火をつけたりすることで、火の体制に影響を与えた最初の一人で あった[206]。火災は、不均質な生態系年齢と樹冠構造を作り出し、変化した土壌栄養供給と除去された樹冠構造は、苗木の定着のための新たな生態学的 ニッチを開く[207][208]。例えば、植物は森林火災に対処するために様々な適応を備えている。いくつかの種(マツ科のマツなど)は、種子が火災を 生き延びたり、煙に含まれる特定の化合物にさらされた後でないと発芽しない。環境によって引き起こされる種子の発芽は、セロティニーと呼ばれる[209] [210]。火は、生態系の持続性と回復力において大きな役割を果たしている[176]。 土壌 主な記事 土壌生態学 土壌とは、地球の表面を覆っている、鉱物や有機物からなる生きた表層のことである。ほとんどの生態系機能の主要な組織化中心であり、農業科学と生態学にお いて極めて重要である。死んだ有機物(例えば林床の葉など)が分解されることで、植物の生産に必要なミネラルや栄養素を含む土壌が形成される。地球の土壌 生態系全体はペドスフィア(土壌圏)と呼ばれ、そこでは地球の生物多様性の大きなバイオマスが栄養レベルに組織化されている。例えば、より大きな葉を食 べ、細断する無脊椎動物は、食物連鎖の中のより小さな生物のために、より小さな断片を作り出す。木の根、菌類、バクテリア、ミミズ、アリ、カブトムシ、ム カデ、クモ、哺乳類、鳥類、爬虫類、両生類、その他あまり馴染みのない生物はすべて、土壌生態系における生命の栄養網を形成するために働いている。土壌 は、無機物が群集全体の生理に包まれた複合表現型を形成している。生物は土壌中で餌を食べたり移動したりする際に、生物撹乱と呼ばれる生態学的プロセスに よって物理的に物質を移動させる。これは生物撹乱と呼ばれる生態学的プロセスであり、土壌を通気し、従属栄養の成長と生産を刺激する。土壌微生物は生態系 の栄養動態の影響を受け、またそれにフィードバックされる。土壌の古生態学的研究では、生物撹乱の起源はカンブリア紀以前の時代とされている[213] [214]。デボン紀における樹木の進化や土地の植民地化など、その他の出来事も、土壌における生態学的トロフィズムの初期の発達に重要な役割を果たした [212][215][216]。 生物地球化学と気候 主な記事 生物地球化学 以下も参照のこと: 栄養塩循環と気候 生態学者は、これらの物質が環境を通してどのように調節され、流れ、リサイクルされているかを理解するために、栄養塩収支を研究・測定している[112] [113][173]。この研究により、生態系と、鉱物、土壌、pH、イオン、水、大気ガスなど、この惑星の物理的パラメータとの間に、グローバルな フィードバックが存在することが理解されるようになった。六大元素(水素、炭素、窒素、酸素、硫黄、リン:H、C、N、O、S、P)は、すべての生物学的 高分子の構成要素であり、地球の地球化学的プロセスに影響を与えている。生物学の最も小さなスケールから、何十億、何百億という生態学的プロセスの複合効 果が増幅され、最終的に地球の生物地球化学サイクルを制御している。これらの元素とその生態学的経路の間に介在する関係とサイクルを理解することは、地球 規模の生物地球化学を理解する上で重要な意味を持つ[217]。 地球規模の炭素収支の生態学は、生物多様性と生物地球化学の関連性の一例を示している。地球の海洋は40,000ギガトン(Gt)の炭素を保有し、植生と 土壌は2070Gtの炭素を保有し、化石燃料の排出量は年間6.3Gtの炭素であると推定されている。例えば、始新世初期から中期にかけての火山噴出物、 湿地帯に貯蔵されたメタンの酸化、海底ガスによって、大気中のCO2(二酸化炭素)濃度は3500ppmという高いレベルまで上昇した[219]。 2,500万年前から3,200万年前の漸新世では、イネ科植物が光合成の新しいメカニズムであるC4光合成を進化させ、その生息域を拡大したため、地球 規模の炭素循環に再び大きな再編成が起こった。この新たな経路は、大気中のCO2濃度が550ppm以下に低下したことに対応して進化したものである。地 球上の生態系に対する人間主導の改変(撹乱、生物多様性の損失、農業など)は、大気中の温室効果ガス濃度の上昇に寄与している。次の世紀における地球規模 の炭素循環の変化は、惑星の気温を上昇させ、天候の極端な変動をもたらし、種の分布を変化させ、絶滅率を増加させると予測されている。地球温暖化の影響 は、氷河の融解、山の氷冠の融解、海面上昇という形ですでに現れている。その結果、移動パターンや繁殖地が気候の変化に追従している水辺や大陸地域で、種 の分布が変化している。永久凍土の大部分も融解し、メタン(CH4)の排出を増加させる土壌分解活動の速度を増加させる、新しいモザイク状の氾濫地域を作 り出している。メタンは温室効果ガスであり、100年の時間スケールで見ると、CO2よりも長波放射を吸収する効果が23倍高いため、地球規模の炭素循環 との関連において大気中のメタンの増加が懸念されている[221]。 |
| History Main article: History of ecology Early beginnings By ecology, we mean the whole science of the relations of the organism to the environment including, in the broad sense, all the "conditions of existence". Thus, the theory of evolution explains the housekeeping relations of organisms mechanistically as the necessary consequences of effectual causes; and so forms the monistic groundwork of ecology. Ernst Haeckel (1866)[227]: 140 [B] Ecology has a complex origin, due in large part to its interdisciplinary nature.[228] Ancient Greek philosophers such as Hippocrates and Aristotle were among the first to record observations on natural history. However, they viewed life in terms of essentialism, where species were conceptualized as static unchanging things while varieties were seen as aberrations of an idealized type. This contrasts against the modern understanding of ecological theory where varieties are viewed as the real phenomena of interest and having a role in the origins of adaptations by means of natural selection.[5][229][230] Early conceptions of ecology, such as a balance and regulation in nature can be traced to Herodotus (died c. 425 BC), who described one of the earliest accounts of mutualism in his observation of "natural dentistry". Basking Nile crocodiles, he noted, would open their mouths to give sandpipers safe access to pluck leeches out, giving nutrition to the sandpiper and oral hygiene for the crocodile.[228] Aristotle was an early influence on the philosophical development of ecology. He and his student Theophrastus made extensive observations on plant and animal migrations, biogeography, physiology, and their behavior, giving an early analogue to the modern concept of an ecological niche.[231][232] Nowhere can one see more clearly illustrated what may be called the sensibility of such an organic complex, – expressed by the fact that whatever affects any species belonging to it, must speedily have its influence of some sort upon the whole assemblage. He will thus be made to see the impossibility of studying any form completely, out of relation to the other forms, – the necessity for taking a comprehensive survey of the whole as a condition to a satisfactory understanding of any part. Stephen Forbes (1887)[233]  Eugenius Warming Ecological concepts such as food chains, population regulation, and productivity were first developed in the 1700s, through the published works of microscopist Antonie van Leeuwenhoek (1632–1723) and botanist Richard Bradley (1688?–1732).[5] Biogeographer Alexander von Humboldt (1769–1859) was an early pioneer in ecological thinking and was among the first to recognize ecological gradients, where species are replaced or altered in form along environmental gradients, such as a cline forming along a rise in elevation. Humboldt drew inspiration from Isaac Newton, as he developed a form of "terrestrial physics". In Newtonian fashion, he brought a scientific exactitude for measurement into natural history and even alluded to concepts that are the foundation of a modern ecological law on species-to-area relationships.[234][235][236] Natural historians, such as Humboldt, James Hutton, and Jean-Baptiste Lamarck (among others) laid the foundations of the modern ecological sciences.[237] The term "ecology" (German: Oekologie, Ökologie) was coined by Ernst Haeckel in his book Generelle Morphologie der Organismen (1866).[238] Haeckel was a zoologist, artist, writer, and later in life a professor of comparative anatomy.[227][239] Opinions differ on who was the founder of modern ecological theory. Some mark Haeckel's definition as the beginning;[240] others say it was Eugenius Warming with the writing of Oecology of Plants: An Introduction to the Study of Plant Communities (1895),[241] or Carl Linnaeus' principles on the economy of nature that matured in the early 18th century.[242][243] Linnaeus founded an early branch of ecology that he called the economy of nature.[242] His works influenced Charles Darwin, who adopted Linnaeus' phrase on the economy or polity of nature in The Origin of Species.[227] Linnaeus was the first to frame the balance of nature as a testable hypothesis. Haeckel, who admired Darwin's work, defined ecology in reference to the economy of nature, which has led some to question whether ecology and the economy of nature are synonymous.[243]  The layout of the first ecological experiment, carried out in a grass garden at Woburn Abbey in 1816, was noted by Charles Darwin in The Origin of Species. The experiment studied the performance of different mixtures of species planted in different kinds of soils.[244][245] From Aristotle until Darwin, the natural world was predominantly considered static and unchanging. Prior to The Origin of Species, there was little appreciation or understanding of the dynamic and reciprocal relations between organisms, their adaptations, and the environment.[229] An exception is the 1789 publication Natural History of Selborne by Gilbert White (1720–1793), considered by some to be one of the earliest texts on ecology.[246] While Charles Darwin is mainly noted for his treatise on evolution,[247] he was one of the founders of soil ecology,[248] and he made note of the first ecological experiment in The Origin of Species.[244] Evolutionary theory changed the way that researchers approached the ecological sciences.[249] Since 1900 Modern ecology is a young science that first attracted substantial scientific attention toward the end of the 19th century (around the same time that evolutionary studies were gaining scientific interest). The scientist Ellen Swallow Richards adopted the term "oekology" (which eventually morphed into home economics) in the U.S. as early as 1892.[250] In the early 20th century, ecology transitioned from a more descriptive form of natural history to a more analytical form of scientific natural history.[234][237][251] Frederic Clements published the first American ecology book in 1905,[252] presenting the idea of plant communities as a superorganism. This publication launched a debate between ecological holism and individualism that lasted until the 1970s. Clements' superorganism concept proposed that ecosystems progress through regular and determined stages of seral development that are analogous to the developmental stages of an organism. The Clementsian paradigm was challenged by Henry Gleason,[253] who stated that ecological communities develop from the unique and coincidental association of individual organisms. This perceptual shift placed the focus back onto the life histories of individual organisms and how this relates to the development of community associations.[254] The Clementsian superorganism theory was an overextended application of an idealistic form of holism.[36][109] The term "holism" was coined in 1926 by Jan Christiaan Smuts, a South African general and polarizing historical figure who was inspired by Clements' superorganism concept.[255][C] Around the same time, Charles Elton pioneered the concept of food chains in his classical book Animal Ecology.[84] Elton[84] defined ecological relations using concepts of food chains, food cycles, and food size, and described numerical relations among different functional groups and their relative abundance. Elton's 'food cycle' was replaced by 'food web' in a subsequent ecological text.[256] Alfred J. Lotka brought in many theoretical concepts applying thermodynamic principles to ecology. In 1942, Raymond Lindeman wrote a landmark paper on the trophic dynamics of ecology, which was published posthumously after initially being rejected for its theoretical emphasis. Trophic dynamics became the foundation for much of the work to follow on energy and material flow through ecosystems. Robert MacArthur advanced mathematical theory, predictions, and tests in ecology in the 1950s, which inspired a resurgent school of theoretical mathematical ecologists.[237][257][258] Ecology also has developed through contributions from other nations, including Russia's Vladimir Vernadsky and his founding of the biosphere concept in the 1920s[259] and Japan's Kinji Imanishi and his concepts of harmony in nature and habitat segregation in the 1950s.[260] Scientific recognition of contributions to ecology from non-English-speaking cultures is hampered by language and translation barriers.[259] This whole chain of poisoning, then, seems to rest on a base of minute plants which must have been the original concentrators. But what of the opposite end of the food chain—the human being who, in probable ignorance of all this sequence of events, has rigged his fishing tackle, caught a string of fish from the waters of Clear Lake, and taken them home to fry for his supper? Rachel Carson (1962)[261]: 48 Ecology surged in popular and scientific interest during the 1960–1970s environmental movement. There are strong historical and scientific ties between ecology, environmental management, and protection.[237] The historical emphasis and poetic naturalistic writings advocating the protection of wild places by notable ecologists in the history of conservation biology, such as Aldo Leopold and Arthur Tansley, have been seen as far removed from urban centres where, it is claimed, the concentration of pollution and environmental degradation is located.[237][262] Palamar (2008)[262] notes an overshadowing by mainstream environmentalism of pioneering women in the early 1900s who fought for urban health ecology (then called euthenics)[250] and brought about changes in environmental legislation. Women such as Ellen Swallow Richards and Julia Lathrop, among others, were precursors to the more popularized environmental movements after the 1950s. In 1962, marine biologist and ecologist Rachel Carson's book Silent Spring helped to mobilize the environmental movement by alerting the public to toxic pesticides, such as DDT, bioaccumulating in the environment. Carson used ecological science to link the release of environmental toxins to human and ecosystem health. Since then, ecologists have worked to bridge their understanding of the degradation of the planet's ecosystems with environmental politics, law, restoration, and natural resources management.[22][237][262][263] |
→「近代生態学の流れ」も参照のこと。 沿革 主な記事 生態学の歴史 初期の始まり 生態学とは、広義にはすべての「存在条件」を含む、生物と環境との関係に関する科学全体を意味する。したがって、進化論は、生物の家事関係を効果的な原因の必然的な結果として機械的に説明し、生態学の一元論的な基礎を形成する。 エルンスト・ヘッケル(1866)[227]: 140 [B] ヒポクラテスやアリストテレスのような古代ギリシャの哲学者たちは、自然史に関する観察を最初に記録したひとりである。しかし彼らは、種は静的で不変のも のとして概念化され、品種は理想化された型の逸脱と見なされていた。このことは、品種が関心のある現実の現象であり、自然淘汰によって適応の起源に関与し ているとみなす現代の生態学理論の理解とは対照的である[5][229][230]。自然における均衡や調整といった生態学の初期の概念は、「自然歯学」 の観察において相互作用の最古の記述のひとつを記したヘロドトス(紀元前425年頃没)にまで遡ることができる。彼は、水浴びをしているナイルワニが口を 開けてイソシギにヒルを安全に吸わせることで、イソシギには栄養を、ワニには口腔衛生を与えていると指摘している。アリストテレスとその弟子テオフラスト スは、動植物の移動、生物地理学、生理学、およびそれらの行動に関する広範な観察を行い、生態学的ニッチという現代の概念に初期の類似性を与えた [231][232]。 このような有機的複合体の感性とでも言うべきものが、これほど明確に示されている場所は他にない。こうして、どの形態も他の形態との関係から切り離して完 全に研究することは不可能であること、つまり、どの部分についても満足のいく理解を得るための条件として、全体を包括的に調査する必要性があることが理解 させられるだろう。 スティーヴン・フォーブス(1887年)[233]。 オイゲニウス・ワーミング 食物連鎖、個体数の調節、生産性といった生態学的概念は、顕微鏡学者アントニー・ファン・レーウェンフック(1632~1723年)や植物学者リチャー ド・ブラッドレー(1688?~1732年)の出版物を通じて、1700年代に初めて発展した[5]。生物地理学者アレクサンダー・フォン・フンボルト (1769~1859年)は、生態学的思考における初期のパイオニアであり、標高の上昇に沿って形成される渓谷のような環境勾配に沿って、種が入れ替わっ たり、形態が変化したりする生態学的勾配を最初に認識したひとりである。フンボルトはアイザック・ニュートンからインスピレーションを受け、「地球物理 学」を発展させた。フンボルト、ジェームズ・ハットン、ジャン=バティスト・ラマルクなどの博物学者は、近代生態学の基礎を築いた。 [237]「生態学」(ドイツ語:Oekologie、Ökologie)という用語は、エルンスト・ヘッケルが著書『Generelle Morphologie der Organismen』(1866年)の中で用いた造語である[238]。ヘッケルは動物学者、芸術家、作家であり、晩年は比較解剖学の教授であった [227][239]。 現代の生態学理論の創始者が誰であるかについては意見が分かれるところである。ヘッケルの定義を始まりとする者もいれば[240]、『植物の生態学』を著 したオイゲニウス・ウォーミングだとする者もいる: 彼の著作はチャールズ・ダーウィンに影響を与え、ダーウィンは『種の起源』において自然の経済または政治に関するリンネの言葉を採用した[227]。ダー ウィンの仕事を賞賛していたヘッケルは、生態学を自然の経済と関連づけて定義しており、そのため生態学と自然の経済が同義語であるかどうかを疑問視する声 もある[243]。 1816年にウォバーン修道院の芝生の庭で行われた最初の生態学的実験のレイアウトは、チャールズ・ダーウィンによって『種の起源』の中で言及された。この実験では、異なる種類の土壌に植えられた異なる種の混合物のパフォーマティビティが研究された[244][245]。 アリストテレスからダーウィンまで、自然界は静的で不変のものと考えられていた。種の起源』以前には、生物とその適応、そして環境との間の動的で相互的な 関係に対する理解や認識はほとんどなかった[229]。例外は1789年に出版されたギルバート・ホワイト(1720-1793)による『セルボーンの博 物誌』であり、生態学に関する最古のテキストのひとつと考えられている。 [チャールズ・ダーウィンは主に進化論で知られているが[247]、土壌生態学の創始者の一人であり[248]、『種の起源』の中で最初の生態学的実験に ついて言及している[244]。 進化論は研究者の生態学への取り組み方を変えた[249]。 1900年以降 現代の生態学は、19世紀末(進化論研究が科学的関心を集めるのと同時期)に初めて科学的注目を集めた若い科学である。科学者エレン・スワロー・リチャー ズは、1892年には早くもアメリカで「エコロジー」(最終的には家庭経済学へと変化した)という用語を採用していた[250]。 20世紀初頭、生態学は自然史のより記述的な形態から科学的自然史のより分析的な形態へと移行した[234][237][251]。フレデリック・クレメ ンツは1905年にアメリカ初の生態学の本を出版し[252]、超組織体としての植物群落の考え方を提示した。この出版は、1970年代まで続いた生態学 的全体主義と個別主義の論争を開始した。クレメンツの超組織体概念は、生態系は生物の発生段階に類似した、鋸歯の発達という規則的で決定された段階を経て 進行すると提唱した。このクレメンツのパラダイムに異議を唱えたのがヘンリー・グリーソンであり[253]、彼は生態系群落は個々の生物のユニークで偶然 の結びつきから発展すると述べた。この認識の転換によって、個々の生物の生活史と、それがどのように群集の連合体の発達に関係しているかに焦点が戻された [254]。 クレメンツのスーパーオーガニズム理論は、ホリズムの理想主義的な形態を過度に拡張したものであった[36][109]。「ホリズム 」という用語は、クレメンツのスーパーオーガニズムの概念に触発された南アフリカの将軍であり、極論的な歴史上の人物であるヤン・クリスティアン・スムッ ツによって1926年に作られた。 [255][C]同じ頃、チャールズ・エルトンは古典的な著書『動物生態学』において食物連鎖の概念を開拓した[84]。エルトン[84]は食物連鎖、食 物循環、食物サイズの概念を用いて生態学的関係を定義し、異なる機能集団間の数値的関係とそれらの相対的な存在量を記述した。エルトンの「食物連鎖」は、 その後の生態学のテキストでは「食物網」に置き換えられた[256]。アルフレッド・J・ロトカは、生態学に熱力学的原理を適用した多くの理論的概念を持 ち込んだ。 1942年、レイモンド・リンデマンは生態学の栄養力学に関する画期的な論文を書いた。栄養動力学は、生態系におけるエネルギーと物質の流れに関する、そ の後の多くの研究の基礎となった。ロバート・マッカーサーは、1950年代に生態学における数学的理論、予測、実験を進め、理論的数理生態学者の復活を促 した。 [237][257][258]生態学はまた、1920年代にロシアのウラジーミル・ヴェルナドスキーと彼の生物圏概念の創設[259]、1950年代に 日本の今西錦司と彼の自然における調和と生息地の隔離の概念[260]など、他の国々からの貢献によっても発展してきた。 英語を話さない文化圏からの生態学への貢献に対する科学的認識は、言語と翻訳の障壁によって妨げられている。 毒の連鎖全体は、最初の濃縮装置であったに違いない微細な植物の基盤の上に成り立っているように思われる。しかし、食物連鎖の対極にある人間はどうだろ う。このような一連の出来事のすべてをおそらく知らないまま、釣り道具に仕掛けをし、クリア湖の水から魚を次々と釣り上げ、家に持ち帰ってフライにして夕 食にするのだろうか。 レイチェル・カーソン(1962)[261]: 48 1960年代から1970年代にかけての環境保護運動において、エコロジーは大衆的かつ科学的な関心を集めた。アルド・レオポルドやアーサー・タンスレー など、保全生物学の歴史において著名な生態学者たちによる、野生の場所の保護を提唱する歴史的強調や詩的自然主義的著作は、汚染や環境悪化の集中が見られ ると主張される都市中心部とはかけ離れたものとみなされてきた[237]。 [237] [262] Palamar(2008)[262]は、都市の健康生態学(当時はユーテニックスと呼ばれた)[250]のために戦い、環境法制に変化をもたらした 1900年代初頭の先駆的な女性たちが、主流派の環境主義によって影が薄くなっていることを指摘している。特にエレン・スワロー・リチャーズやジュリア・ ラスロップといった女性たちは、1950年代以降により一般化した環境運動の先駆者であった。 1962年、海洋生物学者で生態学者でもあるレイチェル・カーソンの著書『沈黙の春』は、DDTなどの有毒農薬が環境中で生物濃縮していることを世間に警 告し、環境保護運動の動員を助けた。カーソンは生態科学を用いて、環境毒素の放出と人間や生態系の健康を関連づけた。それ以来、生態学者は地球の生態系の 劣化に関する理解を、環境政治、法律、修復、自然資源管理と橋渡しすることに取り組んできた[22][237][262][263]。 |
| Carrying capacity Chemical ecology Climate justice Circles of Sustainability Cultural ecology Dialectical naturalism Ecological death Ecological empathy Ecological overshoot Ecological psychology Ecology movement Ecosophy Ecopsychology Human ecology Industrial ecology Information ecology Landscape ecology Natural resource Normative science Philosophy of ecology Political ecology Theoretical ecology Sensory ecology Sexecology Spiritual ecology Sustainable development Lists Glossary of ecology Index of biology articles List of ecologists Outline of biology Terminology of ecology |
環境収容力 ケミカルエコロジー 気候正義 持続可能性の輪 文化的エコロジー 弁証法的自然主義 生態学的死 生態学的共感 生態学的オーバーシュート 生態学的心理学 エコロジー運動 生態哲学 生態心理学 ヒューマンエコロジー 産業生態学 情報生態学 景観生態学 天然資源 規範科学 生態学の哲学 政治生態学 理論生態学 感覚生態学 性生態学 スピリチュアル・エコロジー 持続可能な開発 リスト エコロジー用語集 生物学の記事索引 生態学者のリスト 生物学の概要 生態学の用語 |
| https://en.wikipedia.org/wiki/Ecology |
|
リ ンク
文 献
そ の他の情報
Copyleft, CC, Mitzub'ixi Quq Chi'j, 1996-2099
![]()
☆
 ☆
☆