現代人の近年のアフリカ起源説
Recent African origin of modern humans, OOA
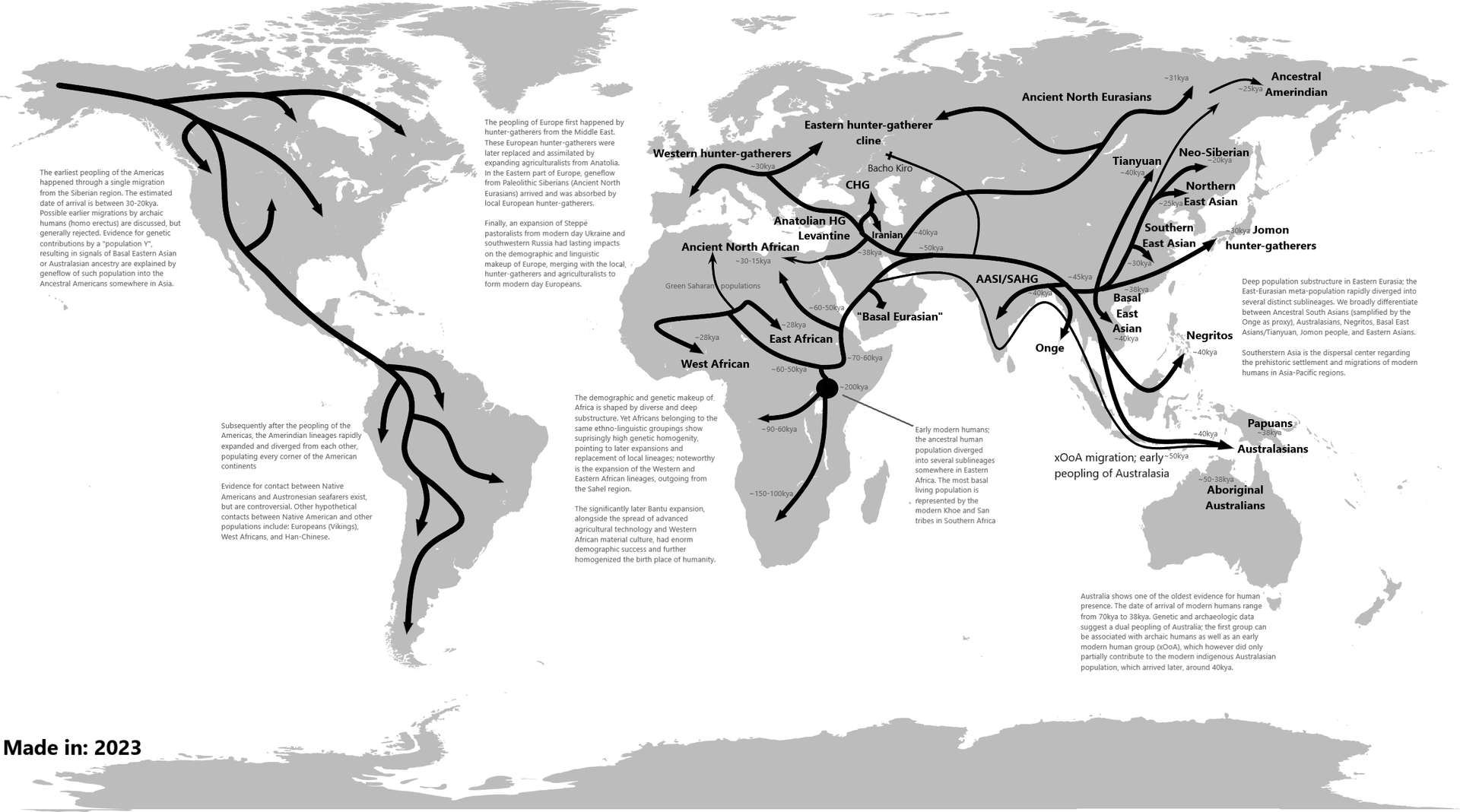
☆ 古人類学研究において、現生人類の最近のアフリカ起源説、または「OOA(Out of Africa)」説は、解剖学的に現生人類(ホモ・サピエンス)の地理的起源と初期の移動に関する最も広く受け入れられているモ デルである。これは、ホモ・エレクトス、そしてホモ・ネアンデルターレンシスによって達成された、アフリカからの 初期の人類の拡散に続くものである。 このモデルは、解剖学的に現生人類とされる特徴が他の地域でも並行して進化していた可能性を排除するものではないが、ヨーロッパやアジアにおける現生人類 と旧人類との複数の混血を排除するものではない。現生人類は、30万年前から2万年前の間にアフリカの角で誕生した可能性が最も高いが、ただし、別の仮説 では、H. sapiensの多様な形態学的特徴は、アフリカの各地域で局所的に出現し、同じ時期に異なる集団間の遺伝子流動によって収束したと主張している。「最近のアフリカ起源」モデルでは、アフリカ以外のすべての現生人類集団 は、その時期以降にアフリカを離れたH. sapiens集団の子孫であると提案している。 少なくとも数回の「アフリカを出た」現代人の拡散があり、おそらく27万年前には始まっており、21万5千年前には少なくともギリシャに到達していた可能 性がある。また、13万年前から11万5千年前にかけては、北アフリカとアラビア半島を経由して確実に拡散していた。約 8万年前に中国に到達していたという証拠がある。これらの初期の波はほぼすべて絶滅または後退したようであり、アフリカ以外の現代人は、主に約7万~5万 年前の単一の拡張、いわゆる「南ルート」を経由して子孫を残したと考えられている。これらの人類はアジアの海岸沿いに急速に広がり、およそ 6万5000年から5万年前にオーストラリアに到達した。(ただし、一部の研究者はオーストラリアへの到達時期を5万年前とみており、それより早い時期に オーストラリアに人類が到達したという説に疑問を呈している。 一方、オーストラリアへの最初の入植者は、アフリカからの大規模な移住以前の古い波を代表するものであり、必ずしもその地域の後の住民の祖先ではない可能 性があることを示唆している)一方、ヨーロッパには、5万5千年前より前に、初期の分派が定住し、近東とヨーロッパに人々が住み着いていた。 2010年代には、集団遺伝学の研究により、ユーラシア、オセアニア、アフリカにおけるホモ・サピエンスと旧人との間の交雑の証拠が発見された。これによ り、現代の集団は主に初期のホモ・サピエンスに由来するが、より少ない程度ではあるが、旧人の地域変種にも由来することが示された。
★こ れに対する批判的論説は「人類学者たちの出アフリカ強迫神経 症」を参照にしてください(→「現代人の多地域起源説」)。
| In
paleoanthropology, the recent African origin of modern humans
or the "Out of Africa" theory (OOA)[a]
is the most widely accepted[1][2][3] model of the geographic origin and
early migration of anatomically modern humans (Homo sapiens). It
follows the early expansions of hominins out of Africa, accomplished by
Homo erectus and then Homo neanderthalensis. The model proposes a "single origin" of Homo sapiens in the taxonomic sense, precluding parallel evolution in other regions of traits considered anatomically modern,[4] but not precluding multiple admixture between H. sapiens and archaic humans in Europe and Asia.[b][5][6] H. sapiens most likely developed in the Horn of Africa between 300,000 and 200,000 years ago,[7][8] although an alternative hypothesis argues that diverse morphological features of H. sapiens appeared locally in different parts of Africa and converged due to gene flow between different populations within the same period.[9][10] The "recent African origin" model proposes that all modern non-African populations are substantially descended from populations of H. sapiens that left Africa after that time. There were at least several "out-of-Africa" dispersals of modern humans, possibly beginning as early as 270,000 years ago, including 215,000 years ago to at least Greece,[11][12][13] and certainly via northern Africa and the Arabian Peninsula about 130,000 to 115,000 years ago.[20] There is evidence that modern humans had reached China around 80,000 years ago.[21] Practically all of these early waves seem to have gone extinct or retreated back, and present-day humans outside Africa descend mainly from a single expansion about 70,000–50,000 years ago,[22][23][24][7][8][25][26][excessive citations] via the so-called "Southern Route". These humans spread rapidly along the coast of Asia and reached Australia by around 65,000–50,000 years ago,[27][28][c] (though some researchers question the earlier Australian dates and place the arrival of humans there at 50,000 years ago at earliest,[29][30] while others have suggested that these first settlers of Australia may represent an older wave before the more significant out of Africa migration and thus not necessarily be ancestral to the region's later inhabitants[24]) while Europe was populated by an early offshoot which settled the Near East and Europe less than 55,000 years ago.[31][32][33] In the 2010s, studies in population genetics uncovered evidence of interbreeding that occurred between H. sapiens and archaic humans in Eurasia, Oceania and Africa,[34][35][36] indicating that modern population groups, while mostly derived from early H. sapiens, are to a lesser extent also descended from regional variants of archaic humans. |
古
人類学において、現生人類の最近のアフリカ起源説、または「OOA(Out of
Africa)」説[a]は、解剖学的に現生人類(ホモ・サピエンス)の地理的起源と初期の移動に関する最も広く受け入れられている[1][2][3]モ
デルである。このモデルは、ホモ・エレクトスとホモ・ネアンデルターレンシスによって成し遂げられた、アフリカからのホミニンの初期の拡大に従っている。 このモデルは分類学的な意味でのホモ・サピエンスの「単一の起源」を提唱しており、解剖学的に現代人と考えられている形質の他の地域での並行進化を排除し ている[4]が、ヨーロッパやアジアにおけるホモ・サピエンスと古人との複数の混血は排除していない[b][5][6]。しかし別の仮説では、サピエンス の多様な形態学的特徴がアフリカのさまざまな地域で局所的に出現し、同時期の異なる集団間の遺伝子流動によって収束したと主張している[9][10]。 「最近のアフリカ起源」モデルでは、現代のアフリカ以外の集団はすべて、その後にアフリカを離れたサピエンスの集団の実質的な子孫であると提唱している。 現生人類の「アフリカ外」への拡散は少なくとも数回あり、恐らく27万年前から始まり、21万5千年前には少なくともギリシャ[11][12][13]、 そして約13万年から11万5千年前にはアフリカ北部とアラビア半島を経由していた。 [20]現生人類が約8万年前に中国に到達していたという証拠がある[21]。これらの初期の波は実質的にすべて絶滅するか後退したようであり、アフリカ 以外の現生人類は主に、いわゆる「南方ルート」を経由して、約7万~5万年前の単一の拡大から派生している[22][23][24][7][8][25] [26][引用過多]。これらの人類はアジアの沿岸に沿って急速に広がり、約6万5千年前から5万年前までにオーストラリアに到達した[27][28] [c](ただし、一部の研究者はそれ以前のオーストラリアの日付に疑問を呈しており、オーストラリアへの人類の到着を早くても5万年前としている、 [29][30]一方で、オーストラリアへの最初の入植者は、より重要なアフリカからの移住の前の古い波である可能性があり、したがって必ずしもこの地域 の後の住民の祖先ではないことを示唆する研究者もいる[24]。 [31][32][33] 2010年代には、集団遺伝学の研究によって、ユーラシア、オセアニア、アフリカにおいてH.サピエンスと古人類との間で起こった交雑の証拠が発見され [34][35][36]、現代の集団は、そのほとんどが初期のH.サピエンスに由来しているものの、古人類の地域的な変種からもより少ない程度ではある が子孫を残していることが示されている。 |
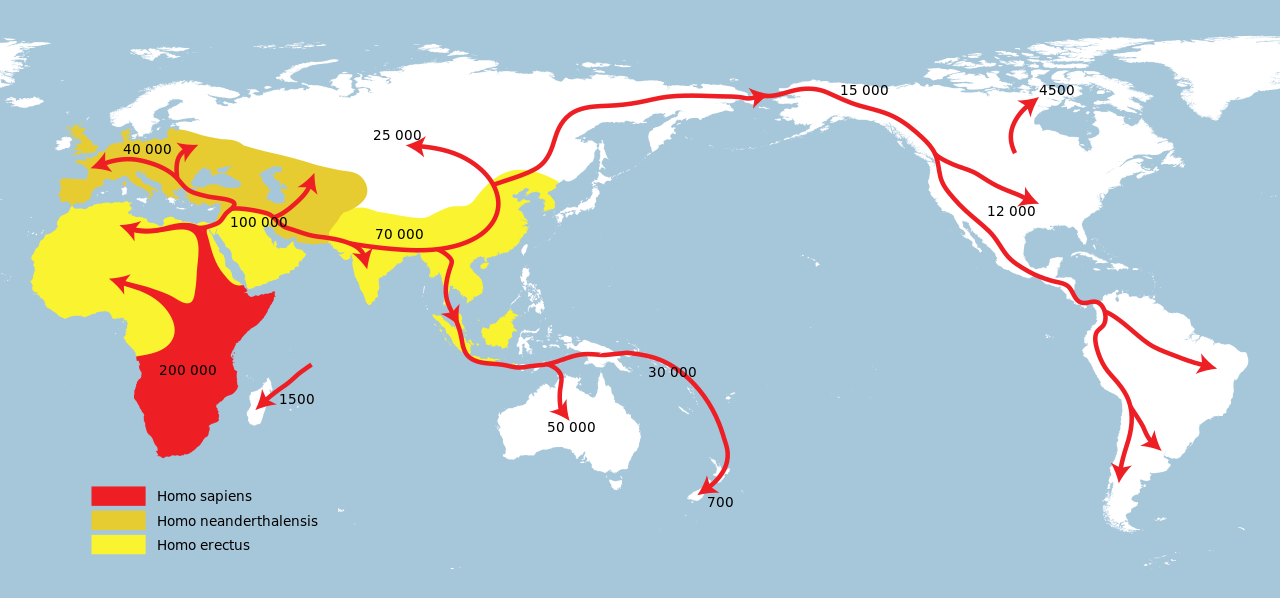
Successive dispersals (labeled in years before present) of Homo erectus greatest extent (yellow) Homo neanderthalensis greatest extent (ochre) Homo sapiens (red) Expansion of early modern humans from Africa through the Near East |
ホモ・エレクトゥスの歴代分散(現在より前の年数で表示)。 ホモ・エレクトスの最大範囲(黄色) ホモ・ネアンデルターレンシス最大範囲(黄土色) ホモ・サピエンス(赤) アフリカから近東を経由した初期現生人類の拡大 |
| Proposed waves Further information: Early expansions of hominins out of Africa and Skhul and Qafzeh hominins 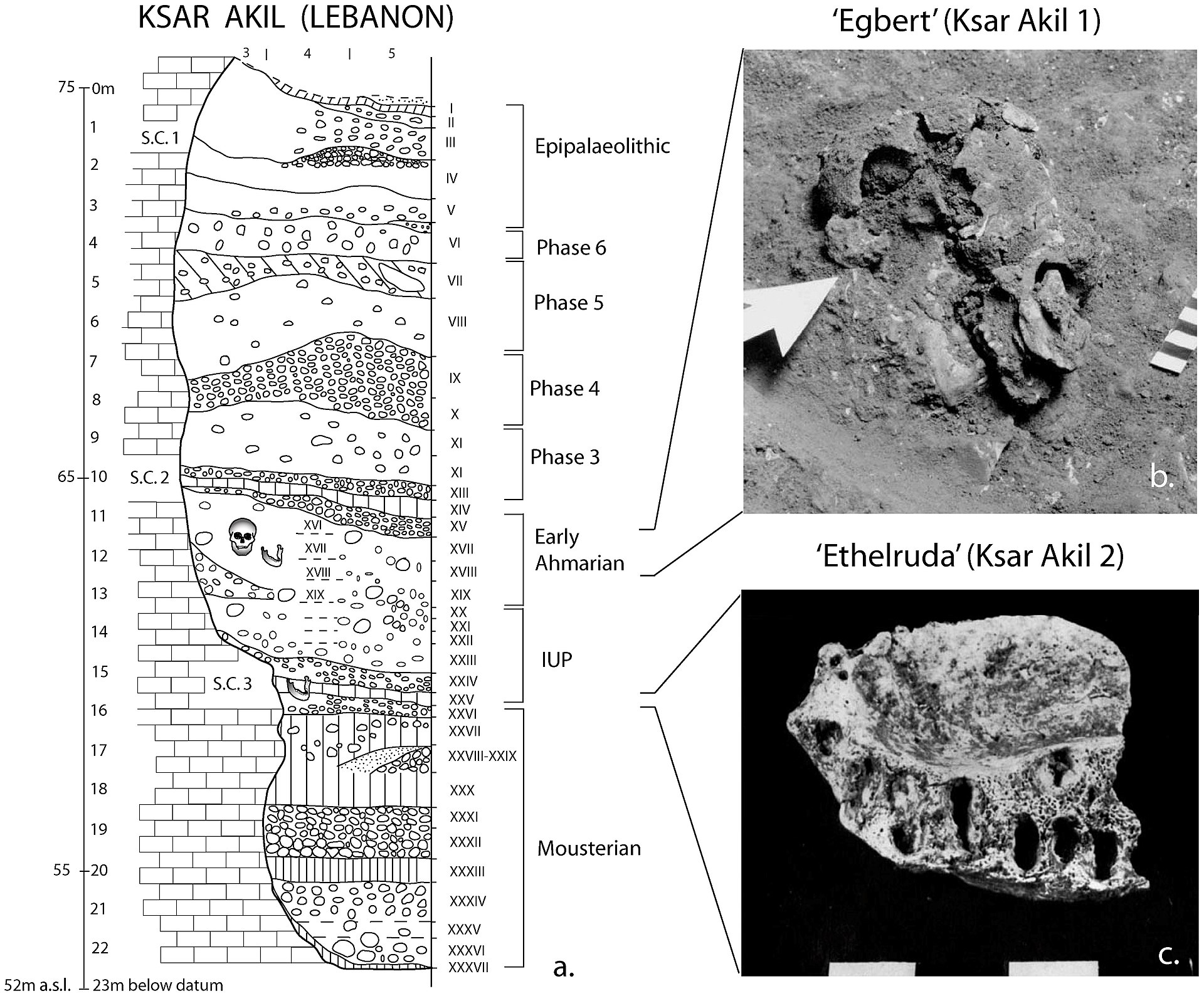 Layer sequence at Ksar Akil in the Levantine corridor, and discovery of two fossils of Homo sapiens, dated to 40,800 to 39,200 years BP for "Egbert",[37] and 42,400–41,700 BP for "Ethelruda".[37] "Recent African origin", or Out of Africa II, refers to the migration of anatomically modern humans (Homo sapiens) out of Africa after their emergence at c. 300,000 to 200,000 years ago, in contrast to "Out of Africa I", which refers to the migration of archaic humans from Africa to Eurasia from before 1.8 and up to 0.5 million years ago. Omo-Kibish I (Omo I) from southern Ethiopia is the oldest anatomically modern Homo sapiens skeleton currently known (around 233,000 years old).[38] There are even older Homo sapiens fossils from Jebel Irhoud in Morocco which exhibit a mixture of modern and archaic features at around 315,000 years old.[39] Since the beginning of the 21st century, the picture of "recent single-origin" migrations has become significantly more complex, due to the discovery of modern-archaic admixture and the increasing evidence that the "recent out-of-Africa" migration took place in waves over a long time. As of 2010, there were two main accepted dispersal routes for the out-of-Africa migration of early anatomically modern humans, the "Northern Route" (via Nile Valley and Sinai) and the "Southern Route" via the Bab-el-Mandeb strait.[40] Posth et al. (2017) suggest that early Homo sapiens, or "another species in Africa closely related to us," might have first migrated out of Africa around 270,000 years ago based on the closer affinity within Neanderthals' mitochondrial genomes to Homo sapiens than Denisovans.[41] Fossil evidence also points to an early Homo sapiens migration with the oldest known fossil coming from Apidima Cave in Greece and dated at 210,000 years ago.[42] Finds at Misliya cave, which include a partial jawbone with eight teeth, have been dated to around 185,000 years ago. Layers dating from between 250,000 and 140,000 years ago in the same cave contained tools of the Levallois type which could put the date of the first migration even earlier if the tools can be associated with the modern human jawbone finds.[43][44][45] An eastward dispersal from Northeast Africa to Arabia 150,000–130,000 years ago is based on the stone tools finds at Jebel Faya dated to 127,000 years ago (discovered in 2011), although fossil evidence in the area is significantly later at 85,000 years ago.[14] [46] Possibly related to this wave are the finds from Zhirendong cave, Southern China, dated to more than 100,000 years ago.[47] Other evidence of modern human presence in China has been dated to 80,000 years ago.[21] The most significant out of Africa dispersal took place around 50,000–70,000 years ago via the so-called Southern Route, either before[48] or after[32][33] the Toba event, which happened between 69,000 and 77,000 years ago.[48] This dispersal followed the southern coastline of Asia and reached Australia around 65,000–50,000 years ago or according to some research, by 50,000 years ago at earliest.[29][30] Western Asia was "re-occupied" by a different derivation from this wave around 50,000 years ago and Europe was populated from Western Asia beginning around 43,000 years ago.[40] Wells (2003) describes an additional wave of migration after the southern coastal route, a northern migration into Europe about 45,000 years ago.[d] This possibility is ruled out by Macaulay et al. (2005) and Posth et al. (2016), who argue for a single coastal dispersal, with an early offshoot into Europe. |
提案された波 さらに詳しい情報 ホミニンのアフリカからの初期拡大、スクールとカフゼのホミニン レヴァント回廊のクサール・アキルで層列が発見され、ホモ・サピエンスの2つの化石が発見されたが、その年代は「エグベルト」が 40,800~39,200年BP[37]、「エセルダ」が42,400~41,700年BPであった[37]。 最近のアフリカ起源」または「アウト・オブ・アフリカII」は、解剖学的に現生人類(ホモ・サピエンス)が30万年から20万年前に出現した後にアフリカ から移動したことを意味し、「アウト・オブ・アフリカI」とは対照的である。エチオピア南部のオモ=キビッシュI(オモI)は、現在知られている中で解剖 学的に最も古い現生ホモ・サピエンスの骨格である(約23万3,000歳)[38]。 モロッコのジェベル・イルフードからは、さらに古いホモ・サピエンスの化石が見つかっている。 21世紀に入ってから、現代人と古人との混血が発見され、「最近の単一起源」移住の図式はかなり複雑になってきている。2010年の時点で、解剖学的に初 期の現生人類のアフリカ外への移動には、「北ルート」(ナイル渓谷とシナイ経由)とバブ・エル・マンデブ海峡経由の「南ルート」の2つの主要な分散ルート が認められている[40]。 Posthら(2017年)は、ネアンデルタール人のミトコンドリアゲノムの中でデニソワ人よりもホモ・サピエンスに近い親和性に基づいて、初期のホモ・ サピエンス、または「私たちに近縁なアフリカの別の種」が、約27万年前にアフリカから最初に移住した可能性を示唆している[41]。 化石の証拠もまた、ギリシャのアピディマ洞窟で発見された最古の化石が21万年前と年代測定されていることから、初期のホモ・サピエンスの移住を指し示し ている[42]。ミスリヤ洞窟で発見された、8本の歯を持つ部分的な顎骨を含む遺物は、約18万5千年前と年代測定されている。同じ洞窟にある25万年か ら14万年前の層には、レヴァロワ型の道具が含まれており、その道具が現生人類の顎骨の出土品と関連づけることができれば、最初の移住の時期をさらに早め ることができる[43][44][45]。 北東アフリカからアラビアへの15万~13万年前の東方への拡散は、ジェベル・ファヤで発見された12万7000年前(2011年に発見された)の石器に 基づくものであるが、この地域における化石の証拠は8万5000年前とかなり後である[14] [46] この波と関連している可能性があるのは、中国南部の志仁洞から発見された10万年以上前のものである[47]。 最も重要なアフリカからの分散は、いわゆる南ルートを通って5万年から7万年前頃に起こったが、これは6万9000年から7万7000年前に起こったト バ・イベントの前[48]か後[32][33]のいずれかであった。 [48]この分散はアジアの南海岸線をたどり、65,000~50,000年前頃に、あるいはいくつかの研究によれば、早くとも50,000年前までには オーストラリアに到達していた[29][30]。西アジアは50,000年前頃にこの波とは異なる派生種によって「再占領」され、ヨーロッパは 43,000年前頃から西アジアから人口が流入していた[40]。 Wells (2003)は、南海岸ルートの後にさらなる移動の波、約45,000年前のヨーロッパへの北方移動を記述している[d]。この可能性は、ヨーロッパへの 早期の分派を伴う単一の海岸拡散を主張するMacaulayら(2005)とPosthら(2016)によって除外されている。 |
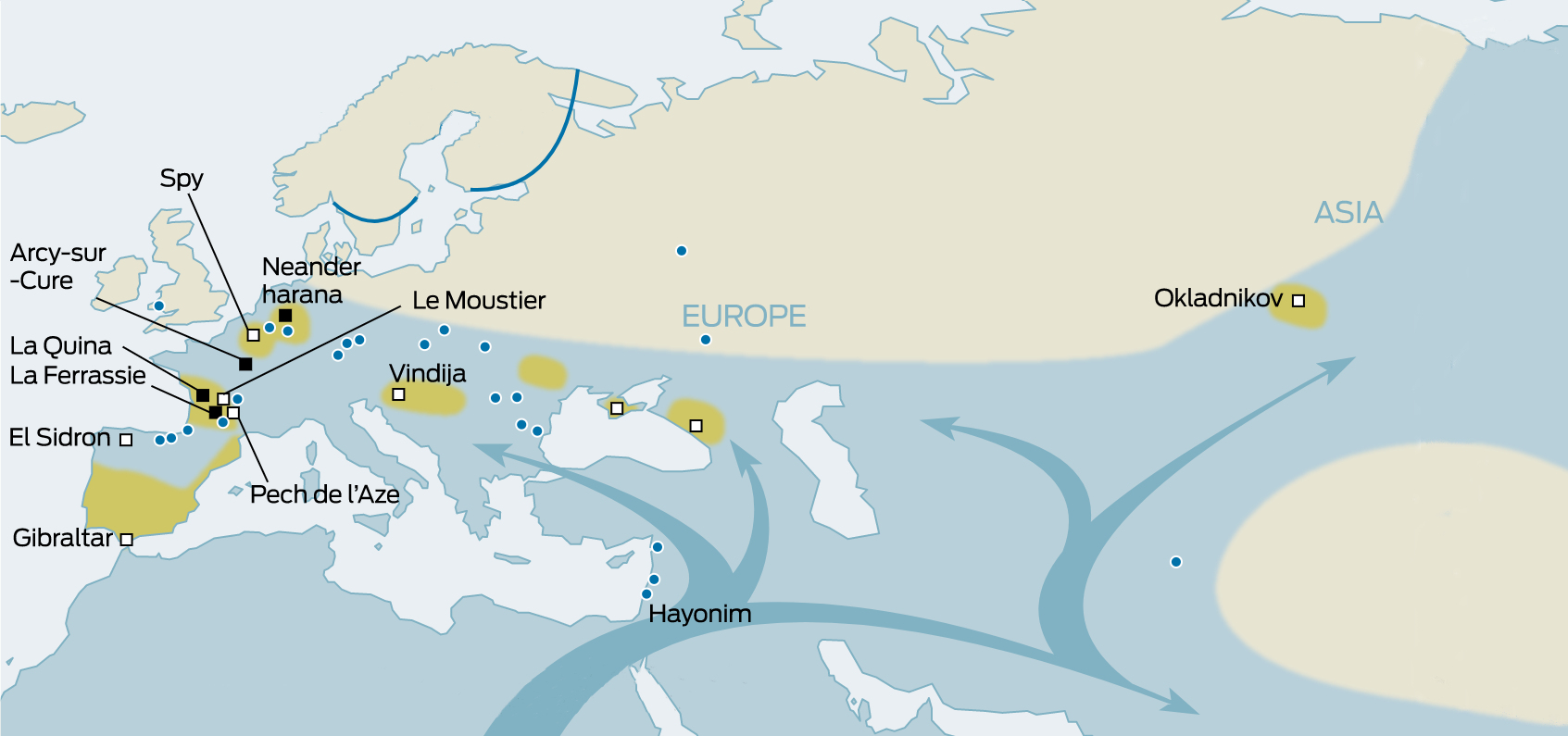
| Northern Route dispersal Further information: Skhul and Qafzeh hominins 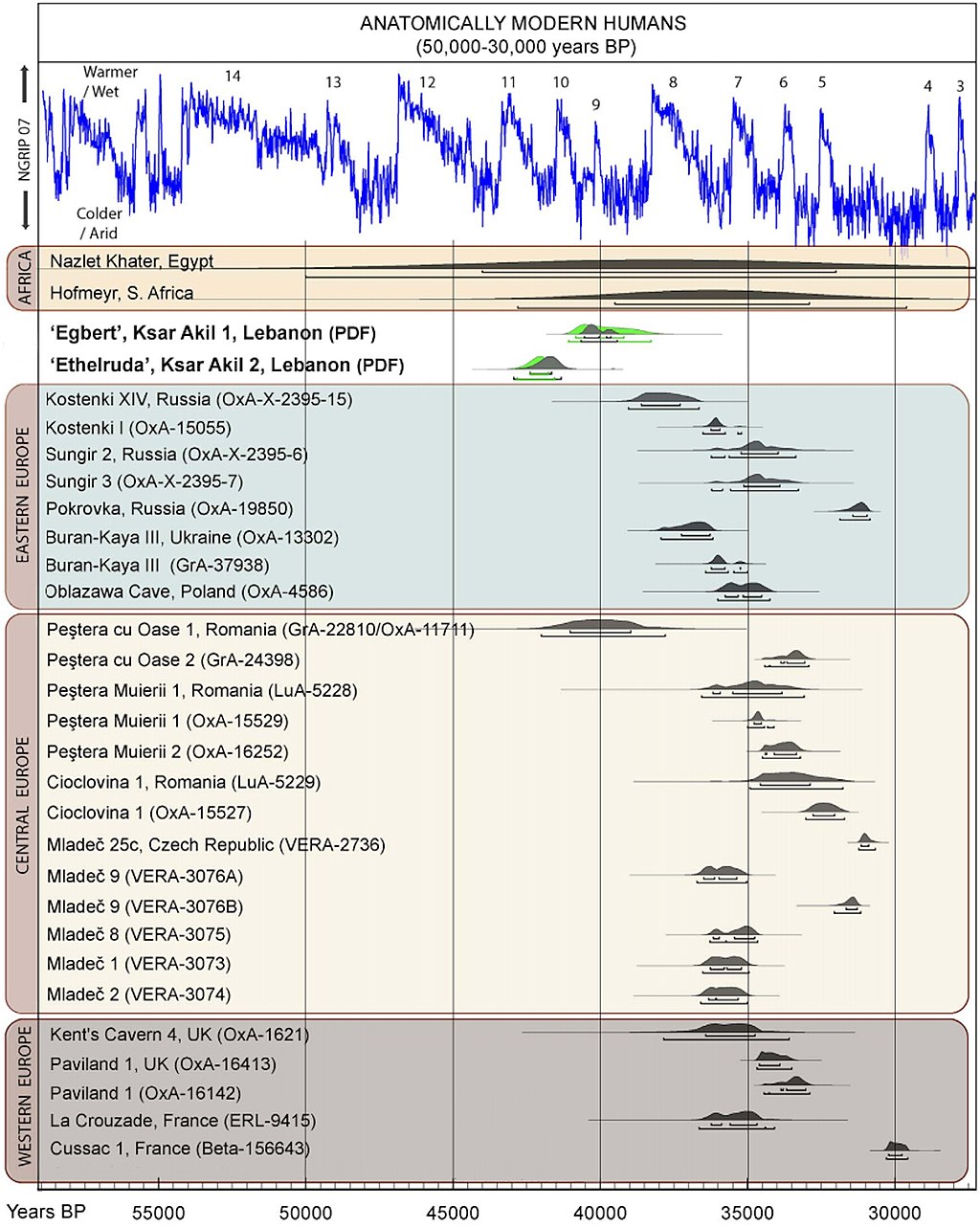 Anatomically Modern Humans known archaeological remains in Europe and Africa, directly dated, calibrated carbon dates as of 2013.[37] Beginning 135,000 years ago, tropical Africa experienced megadroughts which drove humans from the land and towards the sea shores, and forced them to cross over to other continents.[49][e] Fossils of early Homo sapiens were found in Qafzeh and Es-Skhul Caves in Israel and have been dated to 80,000 to 120,000 years ago.[50][51] These humans seem to have either become extinct or retreated back to Africa 70,000 to 80,000 years ago, possibly replaced by southbound Neanderthals escaping the colder regions of ice-age Europe.[22] Hua Liu et al. analyzed autosomal microsatellite markers dating to about 56,000 years ago. They interpret the paleontological fossil as an isolated early offshoot that retracted back to Africa.[23] The discovery of stone tools in the United Arab Emirates in 2011 at the Faya-1 site in Mleiha, Sharjah, indicated the presence of modern humans at least 125,000 years ago,[14] leading to a resurgence of the "long-neglected" North African route.[15][52][16][17] This new understanding of the role of the Arabian dispersal began to change following results from archaeological and genetic studies stressing the importance of southern Arabia as a corridor for human expansions out of Africa.[53] In Oman, a site was discovered by Bien Joven in 2011 containing more than 100 surface scatters of stone tools belonging to the late Nubian Complex, known previously only from archaeological excavations in the Sudan. Two optically stimulated luminescence age estimates placed the Arabian Nubian Complex at approximately 106,000 years old. This provides evidence for a distinct Stone Age technocomplex in southern Arabia, around the earlier part of the Marine Isotope Stage 5.[54] According to Kuhlwilm and his co-authors, Neanderthals contributed genetically to modern humans then living outside of Africa around 100,000 years ago: humans which had already split off from other modern humans around 200,000 years ago, and this early wave of modern humans outside Africa also contributed genetically to the Altai Neanderthals.[55] They found that "the ancestors of Neanderthals from the Altai Mountains and early modern humans met and interbred, possibly in the Near East, many thousands of years earlier than previously thought".[55] According to co-author Ilan Gronau, "This actually complements archaeological evidence of the presence of early modern humans out of Africa around and before 100,000 years ago by providing the first genetic evidence of such populations."[55] Similar genetic admixture events have been noted in other regions as well.[56] |
北ルート分散 さらに詳しい情報 スクル人とカフゼ人 ヨーロッパとアフリカで知られている解剖学的現生人類は、2013年現在、直接年代測定され、較正された炭素年代を持つ考古学的遺跡である[37]。 135,000年前から、熱帯アフリカではメガドライブを経験し、人類は陸地から海辺へと追いやられ、他の大陸へと渡ることを余儀なくされた[49] [e]。 初期のホモ・サピエンスの化石は、イスラエルのカフゼ洞窟とエス・スクル洞窟で発見され、8万年から12万年前と年代測定されている[50][51]。こ れらの人類は、7万年から8万年前に絶滅したか、アフリカに後退したようであり、おそらく氷河期のヨーロッパの寒い地域を逃れて南下したネアンデルタール 人と入れ替わったのであろう。彼らはこの古生物学的化石を、アフリカに後退した孤立した初期の分派であると解釈している[23]。 2011年にアラブ首長国連邦のシャルジャのムレイハにあるファヤ-1遺跡で石器が発見され、少なくとも125,000年前に現生人類が存在していたこと が示され[14]、「長い間無視されてきた」北アフリカのルートが復活するきっかけとなった[15][52][16][17]。アラビア人の分散の役割に 関するこの新しい理解は、アフリカから人類が拡大するための通路としての南アラビアの重要性を強調する考古学的・遺伝学的研究の結果を受けて変わり始めた [53]。 オマーンでは、2011年にビエン・ジョヴェンによって、スーダンの考古学的発掘からのみ知られていた後期ヌビア複合体に属する石器の100以上の表面散 乱を含む遺跡が発見された。2つの光刺激ルミネッセンス法による年代推定では、アラビアン・ヌビア複合体は約10万6000年前のものとされた。このこと は、海洋同位体ステージ5の初期頃に、アラビア南部で明確な石器時代のテクノコンプレックスがあったことを示す証拠となる[54]。 クウルウィルムと彼の共著者たちによれば、ネアンデルタール人は10万年前頃にアフリカの外に住んでいた現生人類に遺伝的に貢献した。アフリカの外にいた 現生人類のこの初期の波は、20万年前頃にすでに他の現生人類から分かれており、アルタイのネアンデルタール人にも遺伝的に貢献した。 [55]彼らは、「アルタイ山脈のネアンデルタール人の祖先と初期現生人類が、おそらくはこれまで考えられていたよりも何千年も早く、近東で出会い、交雑 していた」ことを発見した[55]。共著者のイラン・グロナウによれば、「これは実際に、そのような集団の最初の遺伝的証拠を提供することで、10万年前 以前とその前後にアフリカの外に初期現生人類が存在していたという考古学的証拠を補完するものである」[55]。 |
| Southern Route dispersal Main article: Southern Dispersal Further information: List of first human settlements Coastal route 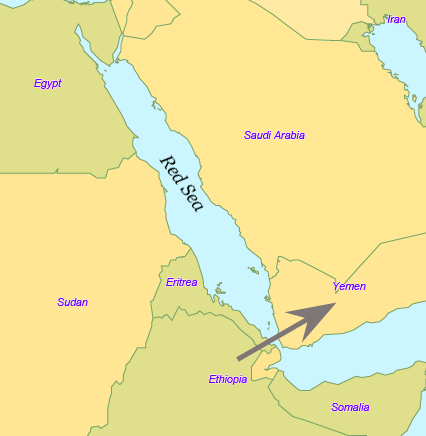 Red Sea crossing By some 50–70,000 years ago, a subset of the bearers of mitochondrial haplogroup L3 migrated from East Africa into the Near East. It has been estimated that from a population of 2,000 to 5,000 individuals in Africa, only a small group, possibly as few as 150 to 1,000 people, crossed the Red Sea.[57][58] The group that crossed the Red Sea travelled along the coastal route around Arabia and the Persian Plateau to India, which appears to have been the first major settling point.[59] Wells (2003) argued for the route along the southern coastline of Asia, across about 250 kilometres (155 mi)[dubious – discuss], reaching Australia by around 50,000 years ago. Migration routes of modern humans, showing the northern route populating Western Eurasia, and the southern/coastal route populating Eastern Eurasia. Today at the Bab-el-Mandeb straits, the Red Sea is about 20 kilometres (12 mi) wide, but 50,000 years ago sea levels were 70 m (230 ft) lower (owing to glaciation) and the water channel was much narrower. Though the straits were never completely closed, they were narrow enough to have enabled crossing using simple rafts, and there may have been islands in between.[40][60] Shell middens 125,000 years old have been found in Eritrea,[61] indicating that the diet of early humans included seafood obtained by beachcombing. Toba eruption Main article: Toba catastrophe theory The dating of the Southern Dispersal is a matter of dispute.[48] It may have happened either pre- or post-Toba, a catastrophic volcanic eruption that took place between 69,000 and 77,000 years ago at the site of present-day Lake Toba in Sumatra, Indonesia. Stone tools discovered below the layers of ash deposited in India may point to a pre-Toba dispersal but the source of the tools is disputed.[48] An indication for post-Toba is haplo-group L3, that originated before the dispersal of humans out of Africa and can be dated to 60,000–70,000 years ago, "suggesting that humanity left Africa a few thousand years after Toba".[48] Some research showing slower than expected genetic mutations in human DNA was published in 2012, indicating a revised dating for the migration to between 90,000 and 130,000 years ago.[62] Some more recent research suggests a migration out-of-Africa of around 50,000-65,000 years ago of the ancestors of modern non-African populations, similar to most previous estimates.[24][63][64] Western Asia Following the fossils dating 80,000 to 120,000 years ago from Qafzeh and Es-Skhul Caves in Israel there are no H. sapiens fossils in the Levant until the Manot 1 fossil from Manot Cave in Israel, dated to 54,700 years ago,[65] though the dating was questioned by Groucutt et al. (2015). The lack of fossils and stone tool industries that can be safely associated with modern humans in the Levant has been taken to suggest that modern humans were outcompeted by Neanderthals until around 55,000 years ago, who would have placed a barrier on modern human dispersal out of Africa through the Northern Route.[66][failed verification] Climate reconstructions also support a Southern Route dispersal of modern humans as the Bab-el-Mandeb strait experienced a climate more conductive to human migration than the northern landbridge to the Levant during the major human dispersal out of Africa.[67] Oceania It is thought that Australia was inhabited around 65,000–50,000 years ago. As of 2017, the earliest evidence of humans in Australia is at least 65,000 years old,[27][28] while McChesney stated that ...genetic evidence suggests that a small band with the marker M168 migrated out of Africa along the coasts of the Arabian Peninsula and India, through Indonesia, and reached Australia very early, between 60,000 and 50,000 years ago. This very early migration into Australia is also supported by Rasmussen et al. (2011).[31] Fossils from Lake Mungo, Australia, have been dated to about 42,000 years ago.[68][69] Other fossils from a site called Madjedbebe have been dated to at least 65,000 years ago,[70][71] though some researchers doubt this early estimate and date the Madjedbebe fossils at about 50,000 years ago at the oldest.[29][30] Phylogenetic data suggests that an early Eastern Eurasian (Eastern non-African) meta-population trifurcated somewhere in eastern South Asia, and gave rise to the Australo-Papuans, the Ancient Ancestral South Indians (AASI), as well as East/Southeast Asians, although Papuans may have also received some gene flow from an earlier group (xOoA), around 2%,[72] next to additional archaic admixture in the Sahul region.[73][74] According to one study, Papuans could have either formed from a mixture between an East Eurasian lineage and lineage basal to West and East Asians, or as a sister lineage of East Asians with or without a minor basal OoA or xOoA contribution.[75] A Holocene hunter-gatherer sample (Leang_Panninge) from South Sulawesi was found to be genetically in between East-Eurasians and Australo-Papuans. The sample could be modeled as ~50% Papuan-related and ~50% Basal-East Asian-related (Andamanese Onge or Tianyuan). The authors concluded that Basal-East Asian ancestry was far more widespread and the peopling of Insular Southeast Asia and Oceania was more complex than previously anticipated.[76][77] 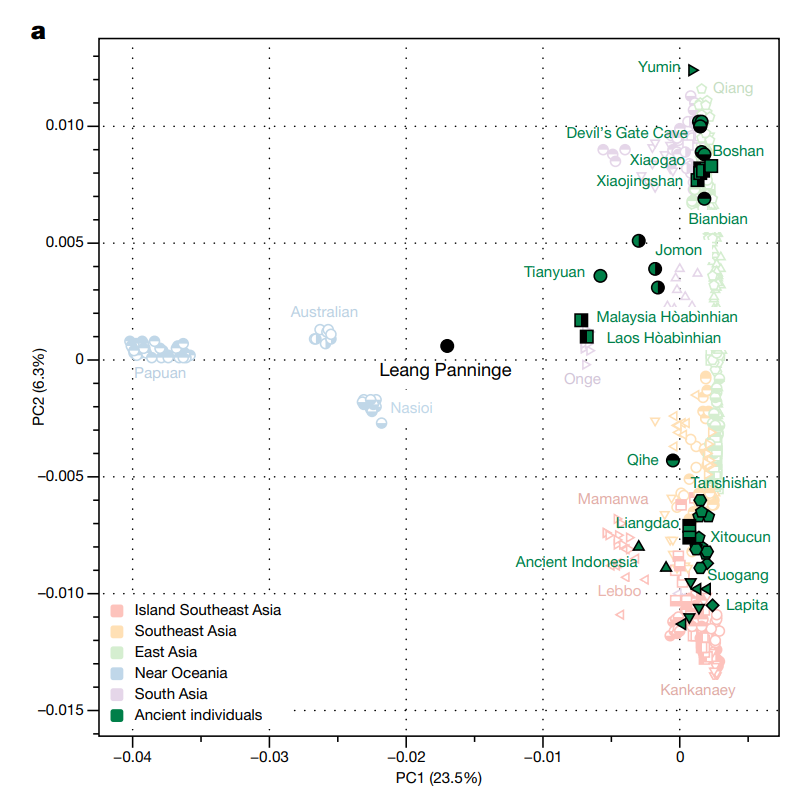 PCA calculated on present-day and ancient individuals from eastern Eurasia and Oceania. PC1 (23,8%) distinguishes East-Eurasians and Australo-Melanesians, while PC2 (6,3%) differentiates East-Eurasians along a North to South cline. 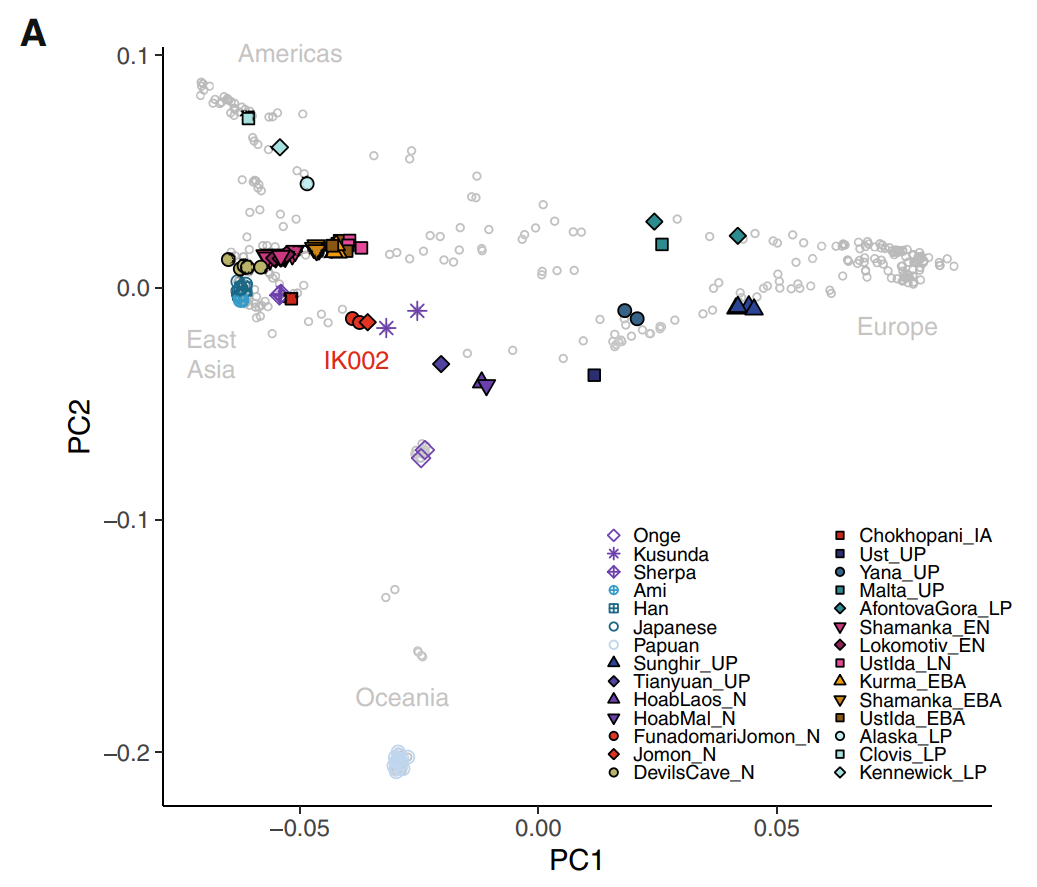 Principal component analysis (PCA) of ancient and modern day individuals from worldwide populations. Oceanians (Aboriginal Australians and Papuans) are most differentiated from both East-Eurasians and West-Eurasians. East and Southeast Asia In China, the Liujiang man (Chinese: 柳江人) is among the earliest modern humans found in East Asia.[78] The date most commonly attributed to the remains is 67,000 years ago.[79] High rates of variability yielded by various dating techniques carried out by different researchers place the most widely accepted range of dates with 67,000 BP as a minimum, but do not rule out dates as old as 159,000 BP.[79] Liu, Martinón-Torres et al. (2015) claim that modern human teeth have been found in China dating to at least 80,000 years ago.[80] Tianyuan man from China has a probable date range between 38,000 and 42,000 years ago, while Liujiang man from the same region has a probable date range between 67,000 and 159,000 years ago. According to 2013 DNA tests, Tianyuan man is related "to many present-day Asians and Native Americans".[81][82][83][84][85] Tianyuan is similar in morphology to Liujiang man, and some Jōmon period modern humans found in Japan, as well as modern East and Southeast Asians.[86][87][88] A 2021 study about the population history of Eastern Eurasia, concluded that distinctive Basal-East Asian (East-Eurasian) ancestry originated in Mainland Southeast Asia at ~50,000BC from a distinct southern Himalayan route, and expanded through multiple migration waves southwards and northwards respectively.[89] Genetic studies concluded that Native Americans descended from a single founding population that initially split from a Basal-East Asian source population in Mainland Southeast Asia around 36,000 years ago, at the same time at which the proper Jōmon people split from Basal-East Asians, either together with Ancestral Native Americans or during a separate expansion wave. They also show that the basal northern and southern Native American branches, to which all other Indigenous peoples belong, diverged around 16,000 years ago.[90][91] An indigenous American sample from 16,000BC in Idaho, which is craniometrically similar to modern Native Americans as well as Paleosiberias, was found to have largely East-Eurasian ancestry and showed high affinity with contemporary East Asians, as well as Jōmon period samples of Japan, confirming that Ancestral Native Americans split from an East-Eurasian source population in Eastern Siberia.[92] Europe According to Macaulay et al. (2005), an early offshoot from the southern dispersal with haplogroup N followed the Nile from East Africa, heading northwards and crossing into Asia through the Sinai. This group then branched, some moving into Europe and others heading east into Asia.[32] This hypothesis is supported by the relatively late date of the arrival of modern humans in Europe as well as by archaeological and DNA evidence.[32] Based on an analysis of 55 human mitochondrial genomes (mtDNAs) of hunter-gatherers, Posth et al. (2016) argue for a "rapid single dispersal of all non-Africans less than 55,000 years ago." By 45,000 years ago, modern humans are known to have reached northwestern Europe.[93] |
南ルート分散 主な記事 南方分散 さらに詳しい情報 人類最初の居住地リスト 沿岸ルート 紅海横断 約5~7万年前までに、ミトコンドリア・ハプログループL3の保持者の一部が東アフリカから近東に移動した。アフリカに2,000人から5,000人いた 集団のうち、紅海を渡ったのはわずかな集団、おそらく150人から1,000人程度であったと推定されている[57][58]。紅海を渡った集団は、アラ ビアとペルシャ高原周辺の海岸沿いのルートを通ってインドまで移動し、そこが最初の主要な定住地であったようである。 [59]ウェルズ(2003年)は、アジアの南海岸線に沿って約250キロメートル(155マイル)を横断するルートを主張し[dubious - discuss]、約5万年前にはオーストラリアに到達していた。 現生人類の移動ルート。西ユーラシアに住む北ルートと、東ユーラシアに住む南/海岸ルートを示す。 今日、バブ・エル・マンデブ海峡の紅海の幅は約20キロメートル(12マイル)だが、5万年前の海面は70メートル(230フィート)低く(氷河期のた め)、水路はもっと狭かった。エリトリアでは12万5,000年前の貝塚が見つかっており[61]、初期の人類の食生活には、浜辺で採れる魚介類が含まれ ていたことを示している[40][60]。 トバ噴火 主な記事 トバ大災害説 南方分散の年代については論争がある[48]。南方分散は、69,000~77,000年前にインドネシアのスマトラ島にある現在のトバ湖で起こった破局 的な火山噴火であるトバ噴火の前後どちらかで起こった可能性がある。インドに堆積した火山灰の層から発見された石器は、トバ以前の飛散を示すかもしれない が、その出所については議論がある[48]。ポスト・トバを示すものとして、ハプログループL3がある。これはアフリカから人類が飛散する前に生まれたも ので、6万~7万年前に年代測定が可能であり、「トバの数千年後に人類がアフリカを離れたことを示唆している」[48]。 [48]。2012年には、ヒトのDNAにおける予想よりも遅い遺伝子変異を示すいくつかの研究が発表され、移住の年代が9万年から13万年前に修正され たことが示された[62]。より最近の研究では、現代の非アフリカ系集団の祖先が約5万年から6万5千年前にアフリカから移住したことを示唆するものもあ り、これはこれまでの推定とほぼ同じである[24][63][64]。 西アジア イスラエルのカフゼ洞窟とエスクル洞窟から発見された80,000年から120,000年前の化石に続き、レバントでは54,700年前とされるイスラエ ルのマノ洞窟のマノ1化石までH.サピエンスの化石は発見されていない[65]。レバントにおける現生人類と安全に関連付けることができる化石や石器産業 の欠如は、現生人類が約5万5千年前までネアンデルタール人によって競争されていたことを示唆しており、ネアンデルタール人は北方ルートを通じてアフリカ から現生人類が分散する際の障壁となっていただろうと考えられている。 オセアニア オーストラリアには約6万5,000~5万年前に人が住んでいたと考えられている。2017年現在、オーストラリアに人類が生息していた最古の証拠は少な くとも6万5,000年前のものであり[27][28]、一方マチェスニーは次のように述べている。 ...遺伝的証拠は、M168というマーカーを持つ小集団がアフリカからアラビア半島とインドの海岸沿いに移動し、インドネシアを経て、6万~5万年前と いう非常に早い時期にオーストラリアに到達したことを示唆している。このオーストラリアへの非常に早い時期の移動は、Rasmussenら(2011年) によっても支持されている[31]。 オーストラリアのマンゴ湖から産出された化石は、約42,000年前と年代測定されている[68][69]。マジェドベベと呼ばれる場所から産出された他 の化石は、少なくとも65,000年前と年代測定されている[70][71]が、一部の研究者はこの早期の推定を疑い、マジェドベベの化石を最も古いもの で約50,000年前と年代測定している[29][30]。 系統発生学的データは、初期の東ユーラシア(東部非アフリカ)のメタ集団が南アジア東部のどこかで三分岐し、オーストラロ・パプア人、古代祖先南インド人 (AASI)、そして東アジア/東南アジア人を生み出したことを示唆しているが、パプア人はサフル地域における古代の混血に次いで、2%程度[72]、そ れ以前のグループ(xOoA)からの遺伝子流入も受けている可能性がある[73][74]。 ある研究によれば、パプア人は東ユーラシア人の系統と西アジア人および東アジア人の基底にある系統との混合から形成されたか、あるいは東アジア人の姉妹系 統として、OoAまたはxOoAの小規模な基底の寄与の有無にかかわらず形成された可能性がある[75]。 南スラウェシの完新世の狩猟採集民サンプル(Leang_Panninge)は、遺伝学的に東ユーラシア人とアウストラロパプア人の中間に位置することが 判明した。このサンプルは、〜50%がパプア人関連で、〜50%が基底東アジア人関連(アンダマン・オンゲまたは天元人)であるとモデル化できた。著者ら は、基底東アジア人の祖先ははるかに広範囲に広がっており、東南アジアとオセアニアの島嶼部の人口形成は以前予想されていたよりも複雑であったと結論づけ た[76][77]。 ユーラシア大陸東部とオセアニアの現生人類と古代の個体について計算されたPCA。PC1(23,8%)は東ユーラシア人とオーストラロメラネシア人を区 別し、PC2(6,3%)は東ユーラシア人を北から南への直線に沿って区別した。 世界人口の古代人と現代人の主成分分析(PCA)。オセアニア人(オーストラリア原住民とパプア人)は、東ユーラシア人および西ユーラシア人の両方と最も 区別される。 東アジアと東南アジア 中国では、柳江人(中国語: 柳江人)が東アジアで発見された最古の現生人類の一人である。 [79]異なる研究者によって実施された様々な年代測定技術によってもたらされた高い変動率は、最も広く受け入れられている年代の範囲を最小値として 67,000BPとするが、159,000BPという古い年代を除外するものではない。[79] Liu, Martinón-Torresら(2015年)は、中国で少なくとも80,000年前の現生人類の歯が発見されたと主張している[80]。 中国の天元人は38,000~42,000年前、同じ地域の柳江人は67,000~159,000年前と推定されている。2013年のDNA検査によれ ば、天元人は「現在のアジア人やアメリカ先住民の多くに近縁」である[81][82][83][84][85]。天元人は、劉江人、日本で発見された縄文 時代の現生人類、現代の東アジア人や東南アジア人と形態が似ている[86][87][88]。 東ユーラシアの人口史に関する2021年の研究では、特徴的な基層東アジア人(東ユーラシア人)の祖先は、ヒマラヤ山脈南部の明確なルートから東南アジア 本土で紀元前5万年頃に発生し、それぞれ南下と北上する複数の移動の波を通して拡大したと結論付けている[89]。 遺伝学的研究は、ネイティブ・アメリカンが東南アジア本土の基底東アジアの源流集団から最初に分かれた単一の創始集団の子孫であり、それは適切な縄文人が 基底東アジア人から分かれたのと同じ時期であり、祖先のネイティブ・アメリカンとともに、あるいは別の拡大波の間に分かれたと結論づけた。また、他のすべ ての先住民族が属している北アメリカ先住民と南アメリカ先住民の分岐は、およそ16,000年前である。 [90][91]アイダホ州の紀元前16,000年前のアメリカ先住民のサンプルは、古オシベリア人と同様に現代のネイティブ・アメリカンと頭蓋測定学的 に類似しているが、大部分が東ユーラシア人の祖先を持っており、現代の東アジア人や日本の縄文時代のサンプルと高い親和性を示していることが発見され、祖 先アメリカ先住民が東シベリアの東ユーラシア人の源集団から分岐したことが確認された[92]。 ヨーロッパ Macaulay et al. (2005)によると、ハプログループNを持つ南方拡散の初期の分派は、東アフリカからナイル川に沿って北上し、シナイ半島を通ってアジアに渡った。この 仮説は、ヨーロッパに現生人類が到着した時期が比較的遅いことや、考古学的証拠やDNAの証拠によって支持されている[32]。狩猟採集民の55のヒトミ トコンドリアゲノム(mtDNA)の分析に基づいて、Posthら(2016)は、「55,000年前未満に、すべての非アフリカ人の急速な単一分散 」を主張している。45,000年前までに、現生人類は北西ヨーロッパに到達したことが知られている[93]。 |
| Genetic reconstruction Mitochondrial haplogroups Within Africa Further information: Most recent common ancestor, Archaeogenetics, and Human mitochondrial DNA haplogroup 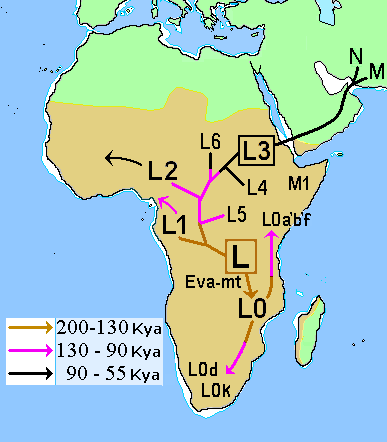 Map of early diversification of modern humans according to mitochondrial population genetics (see: Haplogroup L). The first lineage to branch off from Mitochondrial Eve was L0. This haplogroup is found in high proportions among the San of Southern Africa and the Sandawe of East Africa. It is also found among the Mbuti people.[94][95] These groups branched off early in human history and have remained relatively genetically isolated since then. Haplogroups L1, L2, and L3 are descendants of L1–L6, and are largely confined to Africa. The macro haplogroups M and N, which are the lineages of the rest of the world outside Africa, descend from L3. L3 is about 70,000 years old, while haplogroups M and N are about 65–55,000 years old.[96][64] The relationship between such gene trees and demographic history is still debated when applied to dispersals.[97] Of all the lineages present in Africa, the female descendants of only one lineage, mtDNA haplogroup L3, are found outside Africa. If there had been several migrations, one would expect descendants of more than one lineage to be found. L3's female descendants, the M and N haplogroup lineages, are found in very low frequencies in Africa (although haplogroup M1 populations are very ancient and diversified in North and North-east Africa) and appear to be more recent arrivals.[citation needed] A possible explanation is that these mutations occurred in East Africa shortly before the exodus and became the dominant haplogroups thereafter by means of the founder effect. Alternatively, the mutations may have arisen shortly afterwards. Southern Route and haplogroups M and N Results from mtDNA collected from aboriginal Malaysians called Orang Asli indicate that the haplogroups M and N share characteristics with original African groups from approximately 85,000 years ago, and share characteristics with sub-haplogroups found in coastal south-east Asian regions, such as Australasia, the Indian subcontinent and throughout continental Asia, which had dispersed and separated from their African progenitor approximately 65,000 years ago. This southern coastal dispersal would have occurred before the dispersal through the Levant approximately 45,000 years ago.[32] This hypothesis attempts to explain why haplogroup N is predominant in Europe and why haplogroup M is absent in Europe. Evidence of the coastal migration is thought to have been destroyed by the rise in sea levels during the Holocene epoch.[98] Alternatively, a small European founder population that had expressed haplogroup M and N at first, could have lost haplogroup M through random genetic drift resulting from a bottleneck (i.e. a founder effect). The group that crossed the Red Sea travelled along the coastal route around Arabia and Persia until reaching India.[59] Haplogroup M is found in high frequencies along the southern coastal regions of Pakistan and India and it has the greatest diversity in India, indicating that it is here where the mutation may have occurred.[59] Sixty percent of the Indian population belong to Haplogroup M. The indigenous people of the Andaman Islands also belong to the M lineage. The Andamanese are thought to be offshoots of some of the earliest inhabitants in Asia because of their long isolation from the mainland. They are evidence of the coastal route of early settlers that extends from India to Thailand and Indonesia all the way to eastern New Guinea. Since M is found in high frequencies in highlanders from New Guinea and the Andamanese and New Guineans have dark skin and Afro-textured hair, some scientists think they are all part of the same wave of migrants who departed across the Red Sea ~60,000 years ago in the Great Coastal Migration. The proportion of haplogroup M increases eastwards from Arabia to India; in eastern India, M outnumbers N by a ratio of 3:1. Crossing into Southeast Asia, haplogroup N (mostly in the form of derivatives of its R subclade) reappears as the predominant lineage.[citation needed] M is predominant in East Asia, but amongst Indigenous Australians, N is the more common lineage.[citation needed] This haphazard distribution of Haplogroup N from Europe to Australia can be explained by founder effects and population bottlenecks.[99] 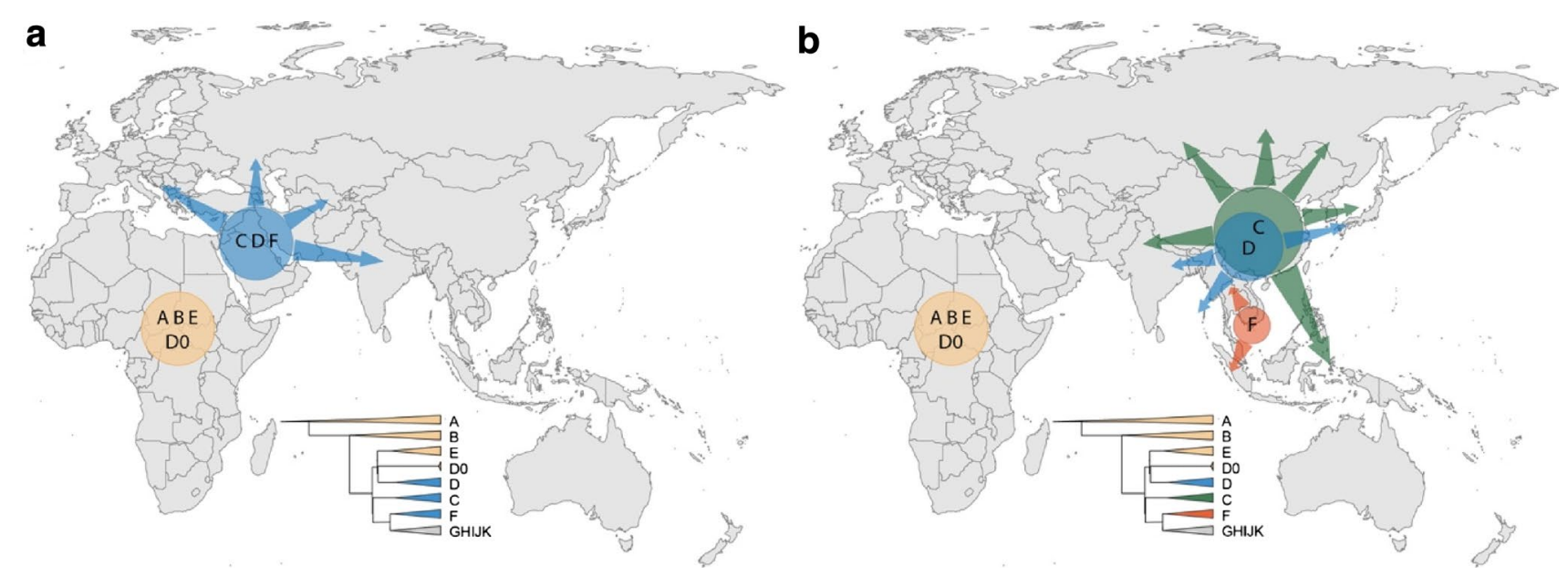 The earliest-branching non-African paternal lineages (C, D, F) after the Out-of-Africa event (a), and their deepest divergence among modern day East or Southeast Asia (b), suggesting rapid coastal expansions. Simplified Y-chromosome tree is shown as reference for colours.[100] Autosomal DNA Further information: Human genetic variation A 2002 study of African, European, and Asian populations, found greater genetic diversity among Africans than among Eurasians, and that genetic diversity among Eurasians is largely a subset of that among Africans, supporting the out of Africa model.[101] A large study by Coop et al. (2009) found evidence for natural selection in autosomal DNA outside of Africa. The study distinguishes non-African sweeps (notably KITLG variants associated with skin color), West-Eurasian sweeps (SLC24A5) and East-Asian sweeps (MC1R, relevant to skin color). Based on this evidence, the study concluded that human populations encountered novel selective pressures as they expanded out of Africa.[102] MC1R and its relation to skin color had already been discussed by Harding et al. (2000), p. 1355. According to this study, Papua New Guineans continued to be exposed to selection for dark skin color so that, although these groups are distinct from Africans in other places, the allele for dark skin color shared by contemporary Africans, Andamanese and New Guineans is an archaism. Endicott et al. (2003) suggest convergent evolution. A 2014 study by Gurdasani et al. indicates that the higher genetic diversity in Africa was further increased in some regions by relatively recent Eurasian migrations affecting parts of Africa.[103] Pathogen DNA Another promising route towards reconstructing human genetic genealogy is via the JC virus (JCV), a type of human polyomavirus which is carried by 70–90 percent of humans and which is usually transmitted vertically, from parents to offspring, suggesting codivergence with human populations. For this reason, JCV has been used as a genetic marker for human evolution and migration.[104] This method does not appear to be reliable for the migration out of Africa; in contrast to human genetics, JCV strains associated with African populations are not basal. From this Shackelton et al. (2006) conclude that either a basal African strain of JCV has become extinct or that the original infection with JCV post-dates the migration from Africa. Admixture of archaic and modern humans Main article: Interbreeding between archaic and modern humans Evidence for archaic human species (descended from Homo heidelbergensis) having interbred with modern humans outside of Africa, was discovered in the 2010s. This concerns primarily Neanderthal admixture in all modern populations except for Sub-Saharan Africans but evidence has also been presented for Denisova hominin admixture in Australasia (i.e. in Melanesians, Aboriginal Australians and some Negritos).[105] The rate of Neanderthal admixture to European and Asian populations as of 2017 has been estimated at between about 2–3%.[106] Archaic admixture in some Sub-Saharan African populations hunter-gatherer groups (Biaka Pygmies and San), derived from archaic hominins that broke away from the modern human lineage around 700,000 years ago, was discovered in 2011. The rate of admixture was estimated at 2%.[36] Admixture from archaic hominins of still earlier divergence times, estimated at 1.2 to 1.3 million years ago, was found in Pygmies, Hadza and five Sandawe in 2012.[107][35] From an analysis of Mucin 7, a highly divergent haplotype that has an estimated coalescence time with other variants around 4.5 million years BP and is specific to African populations, it is inferred to have been derived from interbreeding between African modern and archaic humans.[108] A study published in 2020 found that the Yoruba and Mende populations of West Africa derive between 2% and 19% of their genome from an as-yet unidentified archaic hominin population that likely diverged before the split of modern humans and the ancestors of Neanderthals and Denisovans.[109] Stone tools Further information: Aterian, Baradostian, and Microlith In addition to genetic analysis, Petraglia et al. also examines the small stone tools (microlithic materials) from the Indian subcontinent and explains the expansion of population based on the reconstruction of paleoenvironment. He proposed that the stone tools could be dated to 35 ka in South Asia, and the new technology might be influenced by environmental change and population pressure.[110] |
遺伝的再構築 ミトコンドリアハプログループ アフリカ内 さらに詳しい情報 最近の共通祖先、考古学、ヒト・ミトコンドリアDNAハプログループ ミトコンドリア集団遺伝学による現生人類の初期の多様化の地図(参照:ハプログループL)。 ミトコンドリア・イブから分岐した最初の系統はL0である。このハプログループはアフリカ南部のサン族と東アフリカのサンダウェ族に高い割合で見られる。 これらのグループは人類史の初期に分岐し、それ以来比較的遺伝的に隔離されたままである[94][95]。ハプログループL1、L2、L3はL1-L6の 子孫であり、主にアフリカに限定されている。マクロ・ハプログループMとNは、アフリカ以外の世界の系統であり、L3の子孫である。L3は約7万年前のも のであり、ハプログループMとNは約6万5千年から5万5千年前のものである[96][64]。このような遺伝子系統樹と人口動態史の関係は、分散に当て はめた場合、いまだに議論されている[97]。 アフリカに存在する全系統のうち、女性の子孫はmtDNAハプログループL3という1つの系統だけがアフリカ以外で見つかっている。何度か移動があったの であれば、複数の系統の子孫が見つかると予想される。L3の女性の子孫であるMとNのハプログループ系統は、アフリカでは非常に低い頻度で見つかっており (ただし、ハプログループM1の集団は非常に古く、北アフリカと北東アフリカで多様化している)、最近になって到着したようである。あるいは、突然変異は その直後に発生したのかもしれない。 南ルートとハプログループMとN オラン・アスリと呼ばれるマレーシアの原住民から採取したmtDNAの結果から、ハプログループMとNは、約85,000年前のオリジナルのアフリカ人グ ループと特徴を共有し、オーストラレーシア、インド亜大陸、アジア大陸全域などの東南アジア沿岸地域で発見されたサブハプログループと特徴を共有している ことが示された。この南方沿岸への分散は、約45,000年前にレバントを通って分散する前に起こったと考えられる[32]。この仮説は、ハプログループ Nがヨーロッパで優勢であり、ハプログループMがヨーロッパに存在しない理由を説明しようとするものである。沿岸移動の証拠は完新世の海面上昇によって破 壊されたと考えられている[98]。あるいは、最初にハプログループMとNを発現していたヨーロッパの小さな創始者集団が、ボトルネック(すなわち創始者 効果)の結果生じたランダムな遺伝的ドリフトによってハプログループMを失った可能性もある。 紅海を渡った集団は、アラビアとペルシア周辺の海岸沿いのルートを通ってインドに到達した。ハプログループMはパキスタンとインドの南部沿岸地域に高い頻 度で見られ、インドで最も多様性が高いことから、ここで突然変異が起こった可能性がある。アンダマン諸島の人々は、本土から長い間隔離されていたため、ア ジア最古の住民の子孫であると考えられている。彼らは、インドからタイ、インドネシアを経てニューギニア東部まで続く、初期入植者の沿岸ルートの証拠であ る。Mはニューギニアの高地出身者に高い頻度で見られ、アンダマン人とニューギニア人は黒い肌とアフロ・テクスチャーの髪をしていることから、一部の科学 者は、彼らがすべて、6万年前に紅海を渡って出発した大海岸移動と同じ移民の波の一部であると考えている。ハプログループMの割合は、アラビアからインド に向かって東に増加し、インド東部ではMがNを3:1の割合で上回っている。東南アジアに渡ると、ハプログループN(主にRサブクレードの派生型)が優勢 な系統として再び現れる[要出典]。東アジアではMが優勢であるが、オーストラリア先住民の間ではNの方がより一般的な系統である[要出典]。ヨーロッパ からオーストラリアへのハプログループNのこのような行き当たりばったりの分布は、創始者効果と集団ボトルネックによって説明することができる[99]。 アウト・オブ・アフリカ事象の後に最も早く分岐した非アフリカ父系(C、D、F)(a)、および現代の東アジアまたは東南アジアの間で最も深く分岐してい る(b)は、急速な沿岸部の拡大を示唆している。Y染色体の簡略化されたツリーは、色の参考として示されている[100]。 常染色体DNA さらなる情報 ヒトの遺伝的変異 2002年に行われたアフリカ、ヨーロッパ、アジアの集団の研究では、アフリカ人の遺伝的多様性がユーラシア人の遺伝的多様性よりも大きく、ユーラシア人 の遺伝的多様性はアフリカ人の遺伝的多様性のサブセットであることが発見され、アフリカ外モデルを支持している[101]。この研究では、非アフリカのス イープ(特に肌の色に関連するKITLG変異体)、西ユーラシアのスイープ(SLC24A5)、東アジアのスイープ(肌の色に関連するMC1R)を区別し ている。この証拠に基づいて、この研究は、ヒトの集団がアフリカから拡大する過程で新たな選択圧に遭遇したと結論づけた[102]。MC1Rと肌の色との 関係は、すでにHardingら(2000年)の1355ページで論じられていた。この研究によると、パプアニューギニア人は黒い肌の色の選択にさらされ 続けたため、これらのグループは他の場所のアフリカ人とは異なるが、現代のアフリカ人、アンダマン人、ニューギニア人に共通する黒い肌の色の対立遺伝子は 古風なものである。Endicottら(2003年)は収斂進化を示唆している。Gurdasaniらによる2014年の研究によると、アフリカにおける 遺伝的多様性の高さは、アフリカの一部に影響を与えた比較的最近のユーラシア大陸からの移動によって、いくつかの地域でさらに増加したことが示されている [103]。 病原体DNA ヒトの遺伝的系譜を再構築するためのもう一つの有望なルートは、JCウイルス(JCV)を介したものである。JCVはヒトポリオマウイルスの一種で、 70~90%のヒトが保有しており、通常、親から子へと垂直感染することから、ヒト集団とのコダイバージェンスが示唆されている。このため、JCVはヒト の進化と移動の遺伝子マーカーとして使われてきた。この方法はアフリカからの移動については信頼できないようである。このことから、Shackelton ら(2006年)は、JCVのアフリカの基原株が絶滅したか、あるいはJCVに最初に感染したのがアフリカからの移住より後であったかのどちらかであると 結論づけている。 古人と現代人の混血 主な記事 古人類と現生人類の交雑 古人類(ホモ・ハイデルベルゲンシスの子孫)がアフリカ以外で現生人類と交雑していた証拠が2010年代に発見された。これは主にサハラ以南のアフリカ人 を除くすべての現生人類集団におけるネアンデルタール人との混血に関するものであるが、オーストラレーシア(すなわちメラネシア人、オーストラリアのアボ リジニ、および一部のネグリト人)におけるデニソワ人の混血の証拠も提示されている[105]。2017年現在、ヨーロッパおよびアジアの集団におけるネ アンデルタール人との混血率は約2~3%と推定されている[106]。 サハラ以南のアフリカのいくつかの集団の狩猟採集民グループ(ビアカ・ピグミーとサン)におけるアルカイック混血は、およそ70万年前に現生人類の系統か ら離脱したアルカイック・ホミニンに由来するもので、2011年に発見された。混血率は2%と推定されている[36]。2012年には、120万年から 130万年前と推定される、さらに早い時期に分岐した古人類からの混血がピグミー、ハザ、5人のサンダウェで発見された[107][35]。 ムチン7の分析から、他の変異体との合体時期が450万年BP前後と推定され、アフリカの集団に特異的である高度に分岐したハプロタイプは、アフリカの現 生人類と古生人類の間の交雑に由来すると推測されている[108]。 2020年に発表された研究によると、西アフリカのヨルバとメンデの集団は、ゲノムの2%から19%が、現生人類とネアンデルタール人やデニソワ人の祖先 が分岐する前に分岐した可能性が高い、まだ確認されていない古人類集団に由来していることが判明した[109]。 石器 さらなる情報: アテリアン、バラドスチアン、マイクロリス ペトラグリアらは、遺伝子分析に加えて、インド亜大陸の小型石器(微小石器)を調査し、古環境の復元に基づいて人口の拡大を説明した。彼は、南アジアにお ける石器の年代は35kaとすることができ、新しい技術は環境変化と人口圧力の影響を受けた可能性があると提唱している[110]。 |
| History of the theory Classical paleoanthropology Further information: Timeline of human evolution and Paleoanthropology § History of paleoanthropology 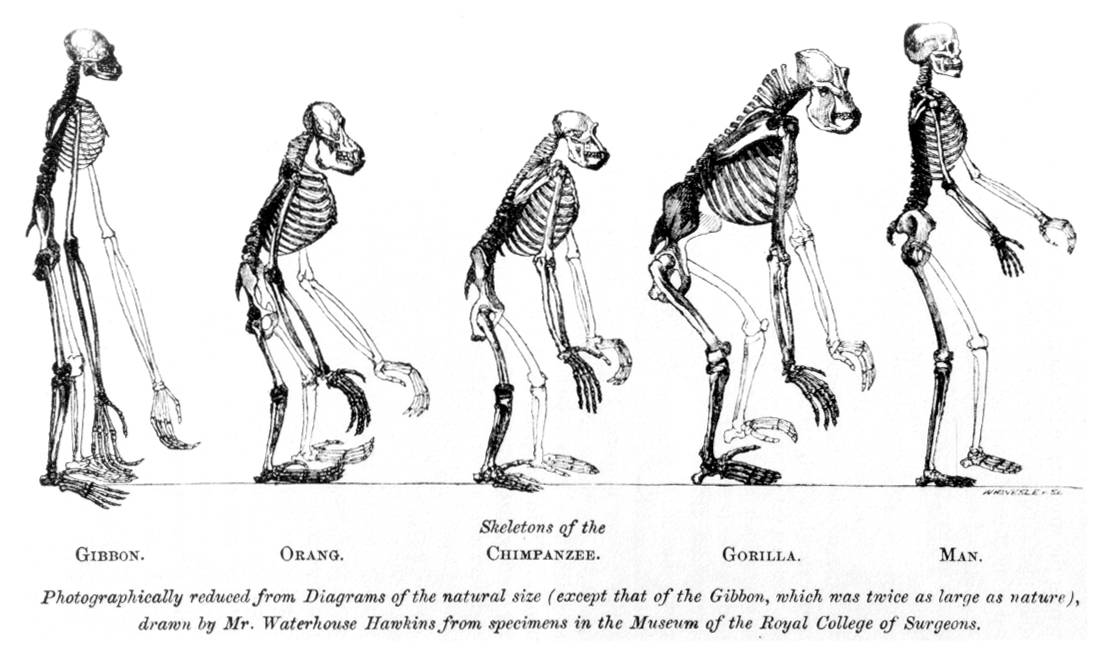 The frontispiece to Huxley's Evidence as to Man's Place in Nature (1863): the image compares the skeleton of a human to other apes. 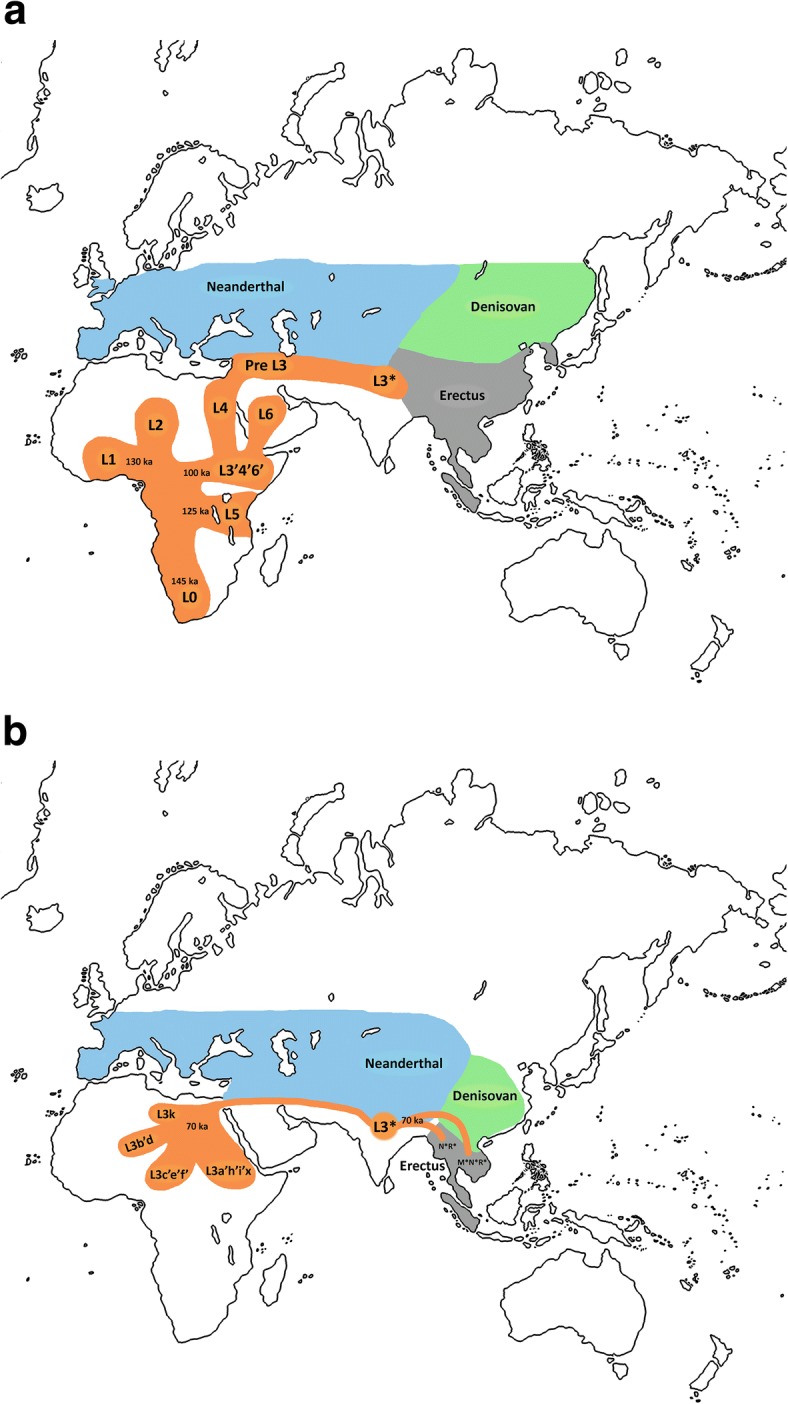 The possibility of an origin of L3 in Asia was proposed by Cabrera et al. (2018).[111] a: Exit of the L3 precursor to Eurasia. b: Return to Africa and expansion to Asia of basal L3 lineages with subsequent differentiation in both continents. The cladistic relationship of humans with the African apes was suggested by Charles Darwin after studying the behaviour of African apes, one of which was displayed at the London Zoo.[112] The anatomist Thomas Huxley had also supported the hypothesis and suggested that African apes have a close evolutionary relationship with humans.[113] These views were opposed by the German biologist Ernst Haeckel, who was a proponent of the Out of Asia theory. Haeckel argued that humans were more closely related to the primates of South-east Asia and rejected Darwin's African hypothesis.[114][115] In The Descent of Man, Darwin speculated that humans had descended from apes, which still had small brains but walked upright, freeing their hands for uses which favoured intelligence; he thought such apes were African: In each great region of the world the living mammals are closely related to the extinct species of the same region. It is, therefore, probable that Africa was formerly inhabited by extinct apes closely allied to the gorilla and chimpanzee; and as these two species are now man's nearest allies, it is somewhat more probable that our early progenitors lived on the African continent than elsewhere. But it is useless to speculate on this subject, for an ape nearly as large as a man, namely the Dryopithecus of Lartet, which was closely allied to the anthropomorphous Hylobates, existed in Europe during the Upper Miocene period; and since so remote a period the earth has certainly undergone many great revolutions, and there has been ample time for migration on the largest scale. — Charles Darwin, Descent of Man[116] In 1871, there were hardly any human fossils of ancient hominins available. Almost fifty years later, Darwin's speculation was supported when anthropologists began finding fossils of ancient small-brained hominins in several areas of Africa (list of hominina fossils). The hypothesis of recent (as opposed to archaic) African origin developed in the 20th century. The "Recent African origin" of modern humans means "single origin" (monogenism) and has been used in various contexts as an antonym to polygenism. The debate in anthropology had swung in favour of monogenism by the mid-20th century. Isolated proponents of polygenism held forth in the mid-20th century, such as Carleton Coon, who thought as late as 1962 that H. sapiens arose five times from H. erectus in five places.[117] Multiregional origin hypothesis Main article: Multiregional origin of modern humans The historical alternative to the recent origin model is the multiregional origin of modern humans, initially proposed by Milford Wolpoff in the 1980s. This view proposes that the derivation of anatomically modern human populations from H. erectus at the beginning of the Pleistocene 1.8 million years BP, has taken place within a continuous world population. The hypothesis necessarily rejects the assumption of an infertility barrier between ancient Eurasian and African populations of Homo. The hypothesis was controversially debated during the late 1980s and the 1990s.[118] The now-current terminology of "recent-origin" and "Out of Africa" became current in the context of this debate in the 1990s.[119] Originally seen as an antithetical alternative to the recent origin model, the multiregional hypothesis in its original "strong" form is obsolete, while its various modified weaker variants have become variants of a view of "recent origin" combined with archaic admixture.[120] Stringer (2014) distinguishes the original or "classic" Multiregional model as having existed from 1984 (its formulation) until 2003, to a "weak" post-2003 variant that has "shifted close to that of the Assimilation Model".[121][122] Mitochondrial analyses Further information: Mitochondrial Eve In the 1980s, Allan Wilson together with Rebecca L. Cann and Mark Stoneking worked on genetic dating of the matrilineal most recent common ancestor of modern human populations (dubbed "Mitochondrial Eve"). To identify informative genetic markers for tracking human evolutionary history, Wilson concentrated on mitochondrial DNA (mtDNA), passed from mother to child. This DNA material mutates quickly, making it easy to plot changes over relatively short times. With his discovery that human mtDNA is genetically much less diverse than chimpanzee mtDNA, Wilson concluded that modern human populations had diverged recently from a single population while older human species such as Neanderthals and Homo erectus had become extinct.[123] With the advent of archaeogenetics in the 1990s, the dating of mitochondrial and Y-chromosomal haplogroups became possible with some confidence. By 1999, estimates ranged around 150,000 years for the mt-MRCA and 60,000 to 70,000 years for the migration out of Africa.[124] From 2000 to 2003, there was controversy about the mitochondrial DNA of "Mungo Man 3" (LM3) and its possible bearing on the multiregional hypothesis. LM3 was found to have more than the expected number of sequence differences when compared to modern human DNA (CRS).[125] Comparison of the mitochondrial DNA with that of ancient and modern aborigines, led to the conclusion that Mungo Man fell outside the range of genetic variation seen in Aboriginal Australians and was used to support the multiregional origin hypothesis. A reanalysis of LM3 and other ancient specimens from the area published in 2016, showed it to be akin to modern Aboriginal Australian sequences, inconsistent with the results of the earlier study.[126] Y-chromosome analyses Further information: Y-chromosomal Adam 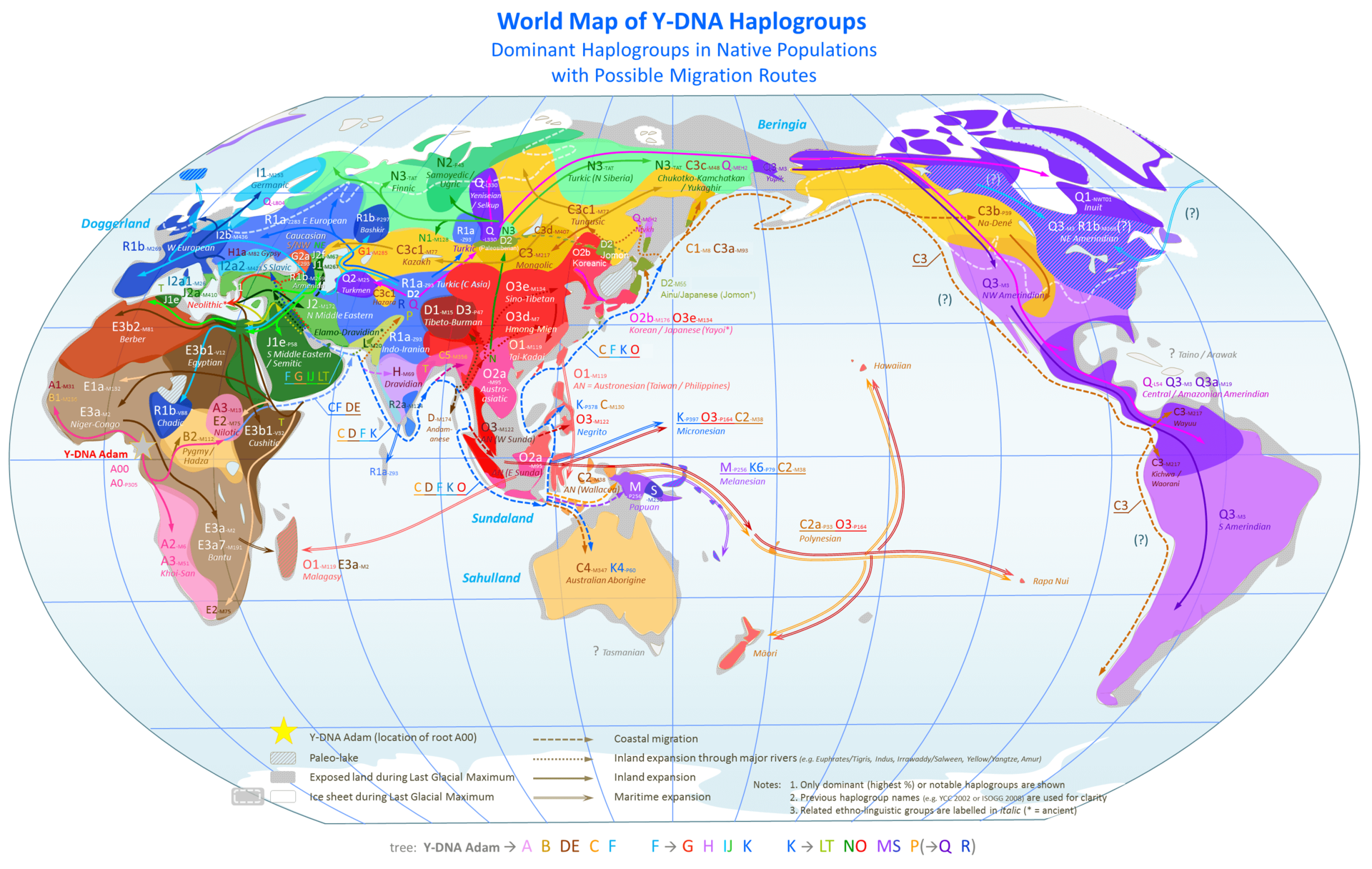 Map of Y-Chromosome Haplogroups – Dominant haplogroups in pre-colonial populations with proposed migrations routes As current estimates on the male most recent common ancestor ("Y-chromosomal Adam" or Y-MRCA) converge with estimates for the age of anatomically modern humans, and well predate the Out of Africa migration, geographical origin hypotheses continue to be limited to the African continent. The most basal lineages have been detected in West, Northwest and Central Africa, suggesting plausibility for the Y-MRCA living in the general region of "Central-Northwest Africa".[16] Another study finds a plausible placement in "the north-western quadrant of the African continent" for the emergence of the A1b haplogroup.[127] The 2013 report of haplogroup A00 found among the Mbo people of western present-day Cameroon is also compatible with this picture.[128] The revision of Y-chromosomal phylogeny since 2011 has affected estimates for the likely geographical origin of Y-MRCA as well as estimates on time depth. By the same reasoning, future discovery of presently-unknown archaic haplogroups in living people would again lead to such revisions. In particular, the possible presence of between 1% and 4% Neanderthal-derived DNA in Eurasian genomes implies that the (unlikely) event of a discovery of a single living Eurasian male exhibiting a Neanderthal patrilineal line would immediately push back T-MRCA ("time to MRCA") to at least twice its current estimate. However, the discovery of a Neanderthal Y-chromosome by Mendez et al. was tempered by a 2016 study that suggests the extinction of Neanderthal patrilineages, as the lineage inferred from the Neanderthal sequence is outside of the range of contemporary human genetic variation.[129] Questions of geographical origin would become part of the debate on Neanderthal evolution from Homo erectus. |
理論の歴史 古典的古人類学 さらなる情報 人類進化の年表と古人類学 § 古人類学の歴史 ハクスリーの『Evidence as to Man's Place in Nature』(1863年)の扉絵:ヒトと他の類人猿の骨格を比較したもの。 アジアにおけるL3の起源の可能性はカブレラら(2018年)によって提唱された[111]。 a: ユーラシア大陸へのL3前駆体の退出。b:アフリカへの帰還と、両大陸でのその後の分化を伴うL3基部系統のアジアへの拡大。 人類とアフリカ類人猿の系統的関係は、チャールズ・ダーウィンが、ロンドン動物園に展示されていたアフリカ類人猿の行動を研究した結果、示唆したものであ る[112]。解剖学者のトーマス・ハクスリーもこの仮説を支持し、アフリカ類人猿が人類と進化的に密接な関係にあることを示唆した[113]。これらの 見解に対して、ドイツの生物学者エルンスト・ヘッケルは反対した。ヘッケルは、ヒトは東南アジアの霊長類により近い関係にあると主張し、ダーウィンのアフ リカ仮説を否定した[114][115]。 ダーウィンは『人間の下降』の中で、人類は類人猿の子孫であり、類人猿はまだ脳が小さいが、直立歩行をし、両手を知性に有利な用途に自由に使えるようにし たと推測し、そのような類人猿はアフリカ人であると考えた: 世界の各大地域では、現生哺乳類は同じ地域の絶滅種と密接な関係がある。したがって、アフリカにはかつてゴリラやチンパンジーに近い絶滅した類人猿が生息 していた可能性が高い。この2種は現在、人間の最も近い盟友であることから、我々の初期の祖先が他の地域よりもアフリカ大陸に住んでいた可能性はいくらか 高い。というのも、人間とほぼ同じ大きさの類人猿、すなわちラルテットのドライオピテクスは、同型類人猿のハイロバテスと密接な関係があり、中新世後期の ヨーロッパに存在していたからである。 - チャールズ・ダーウィン『人間の祖先』[116]。 1871年当時、古代のヒト科動物の化石はほとんどなかった。それから約50年後、人類学者がアフリカのいくつかの地域で古代の小脳のホミニンの化石を発 見し始めたとき、ダーウィンの推測は支持された(ホミニナ化石のリスト)。古代のものではなく)最近のアフリカ起源という仮説は、20世紀に発展した。現 生人類の「最近のアフリカ起源」とは、「単一起源」(単遺伝子主義)を意味し、多遺伝子主義の反意語として様々な文脈で使われてきた。人類学における論争 は、20世紀半ばまでに一元論に傾いた。カールトン・クーンは1962年の時点で、サピエンスは5つの場所でH.エレクトスから5回発生したと考えていた [117]。 多地域起源仮説 主な記事 現生人類の多地域起源 最近の起源モデルに対する歴史的な代替案は、1980年代にミルフォード・ウォルポフによって提唱された現生人類の多地域起源説である。この説は、180 万年前の更新世の初めに、H.エレクトスから解剖学的に現生人類の集団が派生し、連続した世界人口の中で起こったとするものである。この仮説は、ホモの古 代ユーラシア集団とアフリカ集団の間に不妊の壁があったという仮定を必然的に否定するものである。この仮説は1980年代後半から1990年代にかけて論 争となった[118]。現在では「最近の起源」と「アフリカの外」という用語は1990年代にこの論争の文脈で使われるようになった[119]。元々は最 近の起源モデルに対する反語的な代替案とみなされていたが、本来の「強い」形の多地域仮説は時代遅れとなり、その様々な修正された弱い形の仮説は古代の混 血と組み合わされた「最近の起源」という見解の変種となっている。 [120] ストリンガー(2014)は、1984年(その定式化)から2003年まで存在したオリジナルの、あるいは「古典的な」多地域モデルを、2003年以降の 「同化モデルのそれに近く移行した」「弱い」変種と区別している[121][122]。 ミトコンドリア分析 更なる情報 ミトコンドリア・イブ 1980年代、アラン・ウィルソンはレベッカ・L・カンとマーク・ストーンキングと共に、現生人類集団の母系的最新共通祖先(「ミトコンドリア・イブ」と 呼ばれる)の遺伝的年代測定に取り組んだ。人類の進化の歴史を追跡するための有益な遺伝マーカーを特定するため、ウィルソンは母から子へと受け継がれるミ トコンドリアDNA(mtDNA)に注目した。このDNAは突然変異が早く、比較的短期間での変化を簡単にプロットすることができる。ウィルソンは、ヒト のミトコンドリアDNAはチンパンジーのミトコンドリアDNAよりも遺伝学的に多様性が低いことを発見し、ネアンデルタール人やホモ・エレクトスのような 古い人類が絶滅した一方で、現生人類の集団は最近になって単一の集団から分岐したと結論づけた[123]。 1990年代に考古学が登場したことで、ミトコンドリアとY染色体のハプログループの年代測定がある程度確実に行えるようになった。1999年までに、 mt-MRCAについては15万年前後、アフリカからの移住については6万年から7万年前後と推定された[124]。 2000年から2003年にかけて、「マンゴ・マン3」(LM3)のミトコンドリアDNAと多地域仮説との関係について論争が起こった。LM3は現代人の DNA(CRS)と比較したところ、予想以上の数の塩基配列の違いがあることが判明した[125]。ミトコンドリアDNAを古代のアボリジニや現代のアボ リジニのものと比較した結果、マンゴ・マンはオーストラリアのアボリジニに見られる遺伝的変異の範囲外であるという結論に至り、多地域起源仮説を支持する ために使用された。2016年に発表されたLM3とこの地域の他の古代標本の再分析では、LM3は現代のアボリジニ・オーストラリア人の配列に類似してお り、以前の研究結果とは矛盾していることが示された[126]。 Y染色体分析 さらなる情報 Y染色体アダム Y染色体ハプログループの地図 - 植民地以前の集団における優勢なハプログループと推定される移動ルート 現在推定されている男性の最近の共通祖先(「Y染色体アダム」またはY-MRCA)は、解剖学的に現生人類の推定年齢に収束しており、アフリカ大陸からの 移住よりもかなり前であるため、地理的起源仮説はアフリカ大陸に限定され続けている。 最も基本的な系統は西アフリカ、北西アフリカ、中央アフリカで検出されており、Y-MRCAが「中央-北西アフリカ」の一般的な地域に住んでいた可能性を 示唆している[16]。 別の研究では、A1bハプログループの出現が「アフリカ大陸の北西の四分円」に位置することが妥当であるとしている[127]。2013年に現在のカメ ルーン西部のムボ族の間で発見されたハプログループA00の報告も、この図式と一致している[128]。 2011年以降のY染色体系統の改訂は、Y-MRCAの地理的起源と考えられる推定値や、時間の深さに関する推定値に影響を及ぼしている。同じ理由で、現 在知られていない古代のハプログループが現生人類から将来発見された場合、再びこのような修正が行われるであろう。特に、ユーラシア人のゲノム中にネアン デルタール人由来のDNAが1%から4%存在する可能性があるということは、ネアンデルタール人の父系を示す1人の現存するユーラシア人男性が発見された 場合、(可能性は低いが)直ちにT-MRCA(「MRCAまでの時間」)が現在の推定値の少なくとも2倍に押し戻されることを意味する。しかし、メンデス らによるネアンデルタール人のY染色体の発見は、ネアンデルタール人の配列から推測される系統が現代人の遺伝的変異の範囲外であることから、ネアンデル タール人の父系血統の絶滅を示唆する2016年の研究によって緩和された[129]。 |
| Behavioral modernity –
Transition of human species to anthropologically modern behavior Southern Dispersal – Early human migration out of Africa Dawn of Humanity (2015 PBS film) – 2015 American documentary film Early human migrations – Spread of humans from Africa through the world Genetics and archaeogenetics of South Asia – Biological field of study Genetic history of Europe Genetic history of Indigenous peoples of the Americas Genetic history of Italy Genetic history of North Africa – North African genetic history Genetic history of the British Isles Genetic history of the Iberian Peninsula – Ancestry of Spanish and Portuguese people Genetic history of the Middle East Hofmeyr Skull – Hominin fossil Human evolution – Evolutionary process leading to anatomically modern humans Human origins {{Human timeline}} – Hominin events for the last 10 million years Identical ancestors point – Concept in genetic genealogy Indo-Aryan migrations – Migrations of Indo-Aryans into the Indian subcontinent Sahara pump theory – Hypothesis about migration of species between Africa and Eurasia The Incredible Human Journey – British science documentary television series Timeline of human evolution |
行動学的現代性 - 人類学的に現代的な行動への人類種の移行 Southern Dispersal(南方拡散) - 初期の人類のアフリカからの移動 人類の夜明け(2015年PBS映画) - 2015年アメリカのドキュメンタリー映画 初期人類の移動 - アフリカから世界に広がった人類 南アジアの遺伝学と考古学 - 生物学的研究分野 ヨーロッパの遺伝史 アメリカ大陸先住民の遺伝史 イタリアの遺伝史 北アフリカの遺伝史-北アフリカの遺伝史 イギリス諸島の遺伝史 イベリア半島の遺伝史-スペイン人とポルトガル人の祖先 中東の遺伝史 ホフマイヤー頭骨-ヒト科の化石 ヒトの進化 - 解剖学的に現生人類に至る進化の過程 人類の起源 {人類年表}} - 過去1000万年間のヒトの出来事 同一祖先点 - 遺伝系図における概念 インド・アーリア人の移動 - インド亜大陸へのインド・アーリア人の移動 サハラ・ポンプ説 - アフリカとユーラシア間の種の移動に関する仮説 The Incredible Human Journey - イギリスの科学ドキュメンタリー番組シリーズ 人類進化の年表 |
| https://en.wikipedia.org/wiki/Recent_African_origin_of_modern_humans |
|
★こ れに対する批判的論説は「人類学者たちの出アフリカ強迫神経 症」を参照にしてください。
リ ンク
文 献
そ の他の情報
Copyleft, CC, Mitzub'ixi Quq Chi'j, 1996-2099
![]()
☆
 ☆
☆