ネアンデルタール
Neanderthal

Neanderthal man reconstruction, Natural History Museum, London.
ネアンデルタール
Neanderthal
Neanderthal man reconstruction, Natural History Museum, London.
★ネアンデルタール人(Homo neanderthalensis or H. sapiens)は、ユーラシア大陸に約40年まで生息していた絶滅した古人類(一般には別種と考えられているが、ホモ・サピエンスの亜種とみなす人もい る)。sapiens neanderthalensis)は、約4万年前までユーラシア大陸に生息していた絶滅した古人類(一般的には別種と見なされているが、ホモ・サピエン スの亜種と見なす人もいる)である。タイプ標本であるネアンデルタール人は、1856年に現在のドイツのネアンデル渓谷で 発見された(→「古人類と現生人類の交配」「ネアンデルタール絶滅」)。
★以下の記事はウィキペディア英語"Neanderthals"の翻訳(部分)である
| Neanderthals
(/niˈændərˌtɑːl, neɪ-, -ˌθɑːl/ nee-AN-də(r)-TAHL, nay-, -THAHL;[7]
Homo neanderthalensis or H. sapiens neanderthalensis) are an extinct
group of archaic humans (generally regarded as a distinct species,
though some regard it as a subspecies of Homo sapiens) who lived in
Eurasia until about 40,000 years ago.[8][9][10][11] The type specimen,
Neanderthal 1, was found in 1856 in the Neander Valley in present-day
Germany. It is not clear when the line of Neanderthals split from that of modern humans; studies have produced various times ranging from 315,000[12] to more than 800,000 years ago.[13] The date of divergence of Neanderthals from their ancestor H. heidelbergensis is also unclear. The oldest potential Neanderthal bones date to 430,000 years ago, but the classification remains uncertain.[14] Neanderthals are known from numerous fossils, especially from after 130,000 years ago.[15] The reasons for Neanderthal extinction are disputed.[16][17] Theories for their extinction include demographic factors such as small population size and inbreeding, competitive replacement,[18] interbreeding and assimilation with modern humans,[19] change of climate,[20][21][22] disease,[23][24] or a combination of these factors.[22] For much of the early 20th century, European researchers depicted Neanderthals as primitive, unintelligent and brutish. Although knowledge and perception of them has markedly changed since then in the scientific community, the image of the unevolved caveman archetype remains prevalent in popular culture.[25][26] In truth, Neanderthal technology was quite sophisticated. It includes the Mousterian stone-tool industry[27][28] as well as the abilities to create fire,[29][30] build cave hearths[31][32] (to cook food, keep warm, defend themselves from animals, placing it at the centre of their homes),[33] make adhesive birch bark tar,[34] craft at least simple clothes similar to blankets and ponchos,[35] weave,[36] go seafaring through the Mediterranean,[37][38] make use of medicinal plants,[39][40][41] treat severe injuries,[42] store food,[43] and use various cooking techniques such as roasting, boiling,[44] and smoking.[45] Neanderthals consumed a wide array of food, mainly hoofed mammals,[46] but also megafauna,[25][47] plants,[48][49][50] small mammals, birds, and aquatic and marine resources.[51] Although they were probably apex predators, they still competed with cave lions, cave hyenas and other large predators.[52] A number of examples of symbolic thought and Palaeolithic art have been inconclusively[53] attributed to Neanderthals, namely possible ornaments made from bird claws and feathers,[54][55] shells,[56] collections of unusual objects including crystals and fossils,[57] engravings,[58] music production (possibly indicated by the Divje Babe flute),[59] and Spanish cave paintings contentiously[60] dated to before 65,000 years ago.[61][62] Some claims of religious beliefs have been made.[63] Neanderthals were likely capable of speech, possibly articulate, although the complexity of their language is not known.[64][65] Compared with modern humans, Neanderthals had a more robust build and proportionally shorter limbs. Researchers often explain these features as adaptations to conserve heat in a cold climate, but they may also have been adaptations for sprinting in the warmer, forested landscape that Neanderthals often inhabited.[66] They had cold-specific adaptations, such as specialised body-fat storage[67] and an enlarged nose to warm air[68] (although the nose could have been caused by genetic drift[69]). Average Neanderthal men stood around 165 cm (5 ft 5 in) and women 153 cm (5 ft 0 in) tall, similar to pre-industrial modern Europeans.[70] The braincases of Neanderthal men and women averaged about 1,600 cm3 (98 cu in) and 1,300 cm3 (79 cu in), respectively,[71][72][73] which is considerably larger than the modern human average (1,260 cm3 (77 cu in) and 1,130 cm3 (69 cu in), respectively).[74] The Neanderthal skull was more elongated and the brain had smaller parietal lobes[75][76][77] and cerebellum,[78][79] but larger temporal, occipital and orbitofrontal regions.[80][81] The total population of Neanderthals remained low, proliferating weakly harmful gene variants[82] and precluding effective long-distance networks. Despite this, there is evidence of regional cultures and regular communication between communities.[83][84] They may have frequented caves and moved between them seasonally.[85] Neanderthals lived in a high-stress environment with high trauma rates, and about 80% died before the age of 40.[86] The 2010 Neanderthal genome project's draft report presented evidence for interbreeding between Neanderthals and modern humans.[87][88][89] It possibly occurred 316,000 to 219,000 years ago,[90] but more likely 100,000 years ago and again 65,000 years ago.[91] Neanderthals also appear to have interbred with Denisovans, a different group of archaic humans, in Siberia.[92][93] Around 1–4% of genomes of Eurasians, Indigenous Australians, Melanesians, Native Americans and North Africans is of Neanderthal ancestry, while most inhabitants of sub-Saharan Africa have around 0.3% of Neanderthal genes, save possible traces from early sapiens-to-Neanderthal gene flow and/or more recent back-migration of Eurasians to Africa. In all, about 20% of distinctly Neanderthal gene variants survive in modern humans.[94] Although many of the gene variants inherited from Neanderthals may have been detrimental and selected out,[82] Neanderthal introgression appears to have affected the modern human immune system,[95][96][97][98] and is also implicated in several other biological functions and structures,[99] but a large portion appears to be non-coding DNA.[100] https://en.wikipedia.org/wiki/Neanderthal |
ネアンデルタール人(/niˈændərˌtɑ nee-ː,
neɪ-, -ˌθ↪Ll/ nee-AN-də(r)-TAHL, nay-, -THAHL; [7] Homo
neanderthalensis or H.
sapiens)は、ユーラシア大陸に約40年まで生息していた絶滅した古人類(一般には別種と考えられているが、ホモ・サピエンスの亜種とみなす人もい
る)。sapiens
neanderthalensis)は、約4万年前までユーラシア大陸に生息していた絶滅した古人類(一般的には別種と見なされているが、ホモ・サピエン
スの亜種と見なす人もいる)である[8][9][10][11]。タイプ標本であるネアンデルタール人1は、1856年に現在のドイツのネアンデル渓谷で
発見された。 ネアンデルタール人の系統がいつ現生人類の系統から分かれたのかは明らかではない。31万5000年前[12]から80万年以上前[13]まで様々な研究 がなされているが、ネアンデルタール人の祖先であるハイデルベルゲンシス(H. heidelbergensis)からネアンデルタール人が分岐した時期も不明である。最も古いネアンデルタール人の可能性のある骨は43万年前のもので あるが、その分類は不確かなままである[14]。ネアンデルタール人は多くの化石、特に13万年前以降の化石から知られている[15]。 ネアンデルタール人が絶滅した理由については議論があり[16][17]、個体数の少なさや近親交配などの人口学的要因、競争的入れ替わり、[18]現生 人類との交雑や同化、[19]気候の変化、[20][21][22]病気、[23][24]、あるいはこれらの要因の組み合わせなどが考えられている [22]。 20世紀初頭の大部分において、ヨーロッパの研究者たちはネアンデルタール人を原始的で、知性がなく、残忍であると描いていた。それ以来、科学界ではネア ンデルタール人に対する知識や認識は著しく変化したが、進化していない原始人の原型というイメージは大衆文化に広く残っている。モースターの石器産業 [27][28]や火を作り出す能力[29][30]、洞窟に囲炉裏を作る能力[31][32](食べ物を調理し、暖をとり、動物から身を守り、家の中心 に置く)、[33]接着性のある白樺の樹皮のタールを作る能力[34]、 [34]少なくとも毛布やポンチョのような簡単な衣服を作り、[35]織物をし、[36]地中海を航海し、[37][38]薬草を利用し、[39] [40][41]重傷を治療し、[42]食料を貯蔵し、[43]焙煎、煮沸、[44]燻製などの様々な調理法を用いていた。 [45]ネアンデルタール人は、主に有蹄哺乳類[46]だけでなく、巨大哺乳類、[25][47]植物、[48][49][50]小型哺乳類、鳥類、水 生・海洋資源など、幅広い食料を消費していた。 [51]彼らはおそらく頂点捕食者であったが、それでも洞窟ライオン、洞窟ハイエナ、その他の大型捕食者と競合していた。 [すなわち、鳥の爪や羽で作られた装飾品、[54][55]貝殻、[56]水晶や化石を含む珍しい物体のコレクション、[57]彫刻、[58]音楽制作 (おそらくディヴィエ・ベベのフルートによって示された)、[59]スペインの洞窟壁画などである。 [61][62]宗教的な信仰があったという主張もある[63]。 ネアンデルタール人はおそらく言葉を話すことができ、おそらく明瞭であったと思われるが、その言語の複雑さはわかっていない[64][65]。 現生人類と比較すると、ネアンデルタール人はより頑丈な体格をしており、それに比例して手足が短かった。研究者たちはこれらの特徴を寒冷な気候で熱を保存 するための適応であると説明することが多いが、ネアンデルタール人がしばしば生息していた温暖な森林地帯で疾走するための適応であった可能性もある [66]。ネアンデルタール人は特殊な体脂肪の貯蔵[67]や暖かい空気を吸うための鼻の拡大[68]などの寒冷地特有の適応を持っていた(ただし、鼻は 遺伝的なドリフト[69]によって生じた可能性もある)。ネアンデルタール人の平均的な身長は男性で165cm、女性で153cmであり、産業革命前の現 代のヨーロッパ人とほぼ同じであった[70]。ネアンデルタール人の男女の脳容積の平均はそれぞれ約1,600cm3(98立方インチ)、 1,300cm3(79立方インチ)であり[71][72][73]、これは現代人の平均(それぞれ1,260cm3(77立方インチ)、 1,130cm3(69立方インチ))よりもかなり大きい。 [74]ネアンデルタール人の頭蓋骨はより細長く、脳は頭頂葉[75][76][77]と小脳[78][79]は小さかったが、側頭部、後頭部、前頭眼窩 は大きかった[80][81]。 ネアンデルタール人の総人口は少ないままであったため、弱く有害な遺伝子変異が増殖し[82]、効果的な長距離ネットワークが妨げられた。にもかかわら ず、地域文化やコミュニティ間の定期的なコミュニケーションの証拠がある[83][84]。 ネアンデルタール人は外傷率の高い高ストレス環境で生活しており、約80%が40歳までに死亡した[86]。 2010年のネアンデルタール人ゲノムプロジェクトの報告書草案では、ネアンデルタール人と現生人類との交雑の証拠が提示された[87][88] [89]。 316,000年から219,000年前に起こった可能性があるが[90]、10万年前と65,000年前に再び起こった可能性が高い[91]。 ネアンデルタール人はまたシベリアでデニソワ人(古代の人類の別のグループ)と交雑していたようである。 [92][93]ユーラシア人、オーストラリア先住民、メラネシア人、アメリカ先住民、北アフリカ人のゲノムの約1~4%がネアンデルタール人の祖先であ り、サハラ以南のアフリカの住民のほとんどは約0.3%のネアンデルタール人の遺伝子を持っている。ネアンデルタール人から受け継いだ遺伝子変異の多くは 有害で淘汰された可能性があるが[82]、ネアンデルタール人からの導入は現代人の免疫システムに影響を与えているようであり[95][96][97] [98]、その他いくつかの生物学的機能や構造にも関与している[99]。 |
| Neanderthals
( Homo neanderthalensis or Homo sapiens neanderthalensis)[8]
are an
extinct species or subspecies of archaic humans who lived in Eurasia
until about 40,000 years ago.[9][10][11][12] While the "causes of
Neanderthal disappearance about 40,000 years ago remain highly
contested,"[13][14] demographic factors such as small population size,
inbreeding, and random fluctuations are considered probable factors.
Other scholars have proposed competitive replacement,[15] assimilation
into the modern human genome (bred into extinction),[16] great climatic
change,[17][18][19] disease,[20][21] or a combination of these
factors.[19] It is unclear when the line of Neanderthals split from that of modern humans; studies have produced various intervals ranging from 315,000[22] to more than 800,000 years ago.[23] The date of divergence of Neanderthals from their ancestor H. heidelbergensis is also unclear. The oldest potential Neanderthal bones date to 430,000 years ago, but the classification remains uncertain.[24] Neanderthals are known from numerous fossils, especially from after 130,000 years ago.[25] The type specimen, Neanderthal 1, was found in 1856 in the Neander Valley in present-day Germany. For much of the early 20th century, European researchers depicted Neanderthals as primitive, unintelligent, and brutish. Although knowledge and perception of them has markedly changed since then in the scientific community, the image of the unevolved caveman archetype remains prevalent in popular culture.[26][27] Neanderthal technology was quite sophisticated. It includes the Mousterian stone-tool industry[28][29] and ability to create fire[30][31] and build cave hearths,[32][33] make the adhesive birch bark tar,[34] craft at least simple clothes similar to blankets and ponchos,[35] weave,[36] go seafaring through the Mediterranean,[37][38] and make use of medicinal plants,[39][40][41] as well as treat severe injuries,[42] store food,[43] and use various cooking techniques such as roasting, boiling,[44] and smoking.[45] Neanderthals made use of a wide array of food, mainly hoofed mammals,[46] but also other megafauna,[26][47] plants,[48][49][50] small mammals, birds, and aquatic and marine resources.[51] Although they were probably apex predators, they still competed with cave bears, cave lions, cave hyaenas, and other large predators.[52] A number of examples of symbolic thought and Palaeolithic art have been inconclusively[53] attributed to Neanderthals, namely possible ornaments made from bird claws and feathers[54][55] or shells,[56] collections of unusual objects including crystals and fossils,[57] engravings,[58] music production indicated by the Divje Babe flute,[59] and Spanish cave paintings contentiously[60] dated to before 65,000 years ago.[61][62] Some claims of religious beliefs have been made.[63] Neanderthals were likely capable of speech, possibly articulate, although the complexity of their language is not known.[64][65] Compared with modern humans, Neanderthals had a more robust build and proportionally shorter limbs. Researchers often explain these features as adaptations to conserve heat in a cold climate, but they may also have been adaptations for sprinting in the warmer, forested landscape that Neanderthals often inhabited.[66] Nonetheless, they had cold-specific adaptations, such as specialised body-fat storage[67] and an enlarged nose to warm air[68] (although the nose could have been caused by genetic drift[69]). Average Neanderthal men stood around 165 cm (5 ft 5 in) and women 153 cm (5 ft 0 in) tall, similar to pre-industrial modern humans.[70] The braincases of Neanderthal men and women averaged about 1,600 cm3 (98 cu in) and 1,300 cm3 (79 cu in) respectively,[71][72][73] which is within the range of the values for modern humans. The Neanderthal skull was more elongated and had smaller parietal lobes[74][75][76] and cerebellum,[77][78] morphological traits used to assign specimens to species. The total population of Neanderthals remained low, proliferating weakly harmful gene variants,[79] and precluding effective long-distance networks. Despite this, there is evidence of regional cultures and thus of regular communication between communities.[80][81] They may have frequented caves, and moved between them seasonally.[82] Neanderthals lived in a high-stress environment with high trauma rates, and about 80% died before the age of 40.[83] The 2010 Neanderthal genome project's draft report presented evidence for interbreeding between Neanderthals and modern humans.[84][85][86] It possibly occurred 316–219 thousand years ago,[87] but more likely 100,000 years ago and again 65,000 years ago.[88] Neanderthals also appear to have interbred with Denisovans, a different group of archaic humans, in Siberia.[89][90] Around 1–4% of genomes of Eurasians, Australo-Melanesians, Native Americans, and North Africans is of Neanderthal ancestry, while the inhabitants of sub-Saharan Africa have only 0.3% of Neanderthal genes, save possible traces from early Sapiens-to-Neanderthal gene flow and/or more recent back-migration of Eurasians to Africa. In all, about 20% of distinctly Neanderthal gene variants survive today.[91] Although many of the gene variants inherited from Neanderthals may have been detrimental and selected out,[79] Neanderthal introgression appears to have affected the modern human immune system,[92][93][94][95] and is also implicated in several other biological functions and structures,[96] but a large portion appears to be non-coding DNA.[97] https://en.wikipedia.org/wiki/Neanderthal Evolution  It is largely thought that H. heidelbergensis was the last common ancestor of Neanderthals, Denisovans, and modern humans before populations became isolated in Europe, Asia, and Africa, respectively.[137] The taxonomic distinction between H. heidelbergensis and Neanderthals is mostly based on a fossil gap in Europe between 300 and 243 thousand years ago during marine isotope stage 8. "Neanderthals", by convention, are fossils which date to after this gap.[136][26][22] However, 430,000-year old bones at Sima de los Huesos could represent early Neanderthals or a closely related group,[24][138][139] and the 400,000-year old Aroeira 3 could represent a transitional phase. Ancestral and derived morphs could have lived concurrently.[140] It is also possible that there was gene flow between Western Europe and Africa during the Middle Pleistocene, obscuring Neanderthal characteristics in such specimens, namely from Ceprano, Italy, and Sićevo Gorge, Serbia.[24] The fossil record is much more complete from 130,000 years ago onwards,[141] and specimens from this period make up the bulk of known Neanderthal skeletons.[142][143] Dental remains from the Italian Visogliano and Fontana Ranuccio sites indicate that Neanderthal dental features had evolved by around 450–430 thousand years ago during the Middle Pleistocene.[144] There are two main hypotheses regarding the evolution of Neanderthals following the Neanderthal/human split: two-phase and accretion. Two-phase argues that a single major environmental event—such as the Saale glaciation—caused European H. heidelbergensis to increase rapidly in body size and robustness, as well as undergoing a lengthening of the head (phase 1), which then led to other changes in skull anatomy (phase 2).[125] However, Neanderthal anatomy may not have been driven entirely by adapting to cold weather.[66] Accretion holds that Neanderthals slowly evolved over time from the ancestral H. heidelbergensis, divided into four stages: early-pre-Neanderthals (MIS 12, Elster glaciation), pre-Neanderthals sensu lato (MIS 11–9, Holstein interglacial), early Neanderthals (MIS 7–5, Saale glaciation–Eemian), and classic Neanderthals sensu stricto (MIS 4–3, Würm glaciation).[136] Numerous dates for the Neanderthal/human split have been suggested. The date of around 250,000 years ago cites "H. helmei" as being the last common ancestor (LCA), and the split is associated with the Levallois technique of making stone tools. The date of about 400,000 years ago uses H. heidelbergensis as the LCA. Estimates of 600,000 years ago assume that "H. rhodesiensis" was the LCA, which split off into modern human lineage and a Neanderthal/H. heidelbergensis lineage.[145] 800,000 years ago has H. antecessor as the LCA, but different variations of this model would push the date back to 1 million years ago.[24][145] However, a 2020 analysis of H. antecessor enamel proteomes suggests that H. antecessor is related but not a direct ancestor.[146] DNA studies have yielded various results for the Neanderthal/human divergence time, such as 538–315,[22] 553–321,[147] 565–503,[148] 654–475,[145] 690–550,[149] 765–550,[24][89] 741–317,[150] and 800–520 thousand years ago;[151] and a dental analysis concluded before 800,000 years ago.[23] Neanderthals and Denisovans are more closely related to each other than they are to modern humans, meaning the Neanderthal/Denisovan split occurred after their split with modern humans.[24][89][133][152] Assuming a mutation rate of 1 × 10−9 or 0.5 × 10−9 per base pair (bp) per year, the Neanderthal/Denisovan split occurred around either 236–190 or 473–381 thousand years ago respectively.[89] Using 1.1 × 10−8 per generation with a new generation every 29 years, the time is 744,000 years ago. Using 5 × 10−10 nucleotide sites per year, it is 616,000 years ago. Using the latter dates, the split had likely already occurred by the time hominins spread out across Europe, and unique Neanderthal features had begun evolving by 600–500 thousand years ago.[133] Before splitting, Neanderthal/Denisovans (or "Neandersovans") migrating out of Africa into Europe apparently interbred with an unidentified "superarchaic" human species who were already present there; these superarchaics were the descendants of a very early migration out of Africa around 1.9 mya.[153] https://en.wikipedia.org/wiki/Neanderthal |
ネ
アンデルタール人( Homo neanderthalensis or Homo sapiens neanderthalensis)[8]
は絶滅種または亜種の古代の人類で、約4万年前まではユーラシアに居住していた。 [9][10][11][12]
「約4万年前のネアンデルタール人の消滅の原因については依然として非常に論争されている」が[13][14]、小さな集団サイズ、近親交配、ランダムな
変動などの人口学的要因が考えられるとされる。他の学者は、競争的置換、[15]現代人ゲノムへの同化(繁殖による絶滅)、[16]大きな気候変動、
[17][18][19]病気、[20][21]、あるいはこれらの要因の組合せを提案している[19]。 ネアンデルタール人の系統が現代人の系統といつ分かれたのかは不明であり、研究によって315,000年[22]から80万年以上前までの様々な間隔が生 み出されている。 ネアンデルタール人の祖先のハイデルベルゲンから分岐した日付も不明である[23]。ネアンデルタール人の可能性のある最も古い骨は43万年前のものであ るが、分類は不明のままである[24]。 ネアンデルタール人は多くの化石から、特に13万年前以降のものが知られている[25]。タイプ標本のネアンデルタール1が1856年に現在のドイツのネ アンデル谷で発見された。20世紀初頭の大部分において、ヨーロッパの研究者はネアンデルタール人を原始的で、知性がなく、残忍であると描写した。それ以 来、科学界では彼らに対する知識と認識は著しく変化したが、進化していない原始人の原型のイメージは大衆文化に広く残っている[26][27]。 ネアンデルタール人の技術はかなり高度なものであった。これにはモウステリアン石器産業[28][29]や火を起こす能力[30][31]や洞窟の囲炉裏 を作る能力[32][33]、接着剤の白樺皮タールの作成[34]、少なくとも毛布やポンチョに似た簡単な衣類の製造、織物[35]が含まれている。 [地中海を航海し[36]、薬草を利用し[39][40][41]、重傷を治療し、食料を貯蔵し、焙煎、煮沸、燻製などの様々な調理技術を利用した [42]。 [ネアンデルタール人は主に有蹄哺乳類[46]だけでなく他の巨大動物[26][47]、植物、[48][49][50]、小型哺乳類、鳥、水中・海洋資 源など幅広い食料を利用していた。 [51]おそらく頂点捕食者であったが、それでもホラアナグマ、ホラアナライオン、ホラアナハイエナ、その他の大型捕食者と競争していた。 [すなわち、鳥の爪や羽[54][55]や貝殻から作られた装飾品[56]、水晶や化石を含む珍しい物のコレクション[57]、彫刻[58]、ディビエ・ バベ笛に示された音楽制作[59]、65000年前以前とする論争[60]のスペイン石窟壁画などである。 [ネアンデルタール人はおそらく言葉を話すことができ、おそらく明瞭であったが、彼らの言語の複雑さは知られていない[64][65]。 現代人と比較して、ネアンデルタール人はより頑丈な体格と比例して短い手足を持っていた。研究者はこれらの特徴を寒い気候で熱を保存するための適応として 説明することが多いが、それらはネアンデルタール人がしばしば生息していた暖かく森林に覆われた景観で疾走するための適応でもあったかもしれない[66] にもかかわらず、彼らは特別な体脂肪の貯蔵[67]や暖かい空気のために拡大した鼻などの寒冷特異的適応を持っていた[68](ただし鼻は遺伝子ドリフト によって引き起こされたかもしれなかったが[69] )。ネアンデルタール人の平均的な男性の身長は約165cm(5フィート5インチ)、女性は153cm(5フィート0インチ)で、産業革命前の現代人と同 様であった[70]。ネアンデルタール人の男性と女性の脳槽はそれぞれ平均約1,600cm3(98立方インチ)、1,300cm3(79立方インチ) で、現代人の数値と同じ範囲であった[71][72][73]。ネアンデルタール人の頭蓋骨はより細長く、頭頂葉[74][75][76]や小脳が小さ く、標本を種に割り当てるために用いられる形態的特徴[77][78]を持っていた。 ネアンデルタール人の総人口は少ないままであり、弱く有害な遺伝子変異を増殖させ[79]、効果的な長距離のネットワークができないままであった。それに もかかわらず、地域文化の証拠があり、したがってコミュニティ間の定期的なコミュニケーションもあった[80][81]。 [2010年のネアンデルタール人ゲノムプロジェクトのドラフトレポートはネアンデルタール人と現代人の間の交雑の証拠を提示した[84][85] [86] それはおそらく316-219千年前に起こったが[87]、10万年前と再び65千年前の可能性がより高いとされた。 [ネアンデルタール人はまた、シベリアで古代の人間の別のグループであるデニソワ人と交配したようである[89][90] ユーラシア人、オーストラリア-メラネシア人、アメリカ先住民、北アフリカ人のゲノムのおよそ1-4%がネアンデルタールの祖先であり、サハラ以南のアフ リカ住民には、初期のサピエンスからネアンデルタールへの遺伝子流、及び/またはより最近のユーラシア人のアフリカへの回帰移住から考えられる痕跡を残し て、0.3%のネアンデルタールの遺伝子しか有していない。ネアンデルタール人から受け継いだ遺伝子変異の多くは有害であり淘汰されたかもしれないが [79]、ネアンデルタール人の導入は現代人の免疫システムに影響を与えているようであり[92][93][94][95]、その他いくつかの生体機能や 構造にも関係しているが、大部分は非コードDNAと思われる[96]。 進化 ハイデルベルグレンシスは、ヨーロッパ、アジア、アフリカでそれぞれ集団が孤立する前のネアンデルタール人、デニソワ人、現代人の最後の共通祖先であると 主に考えられている[137]。ハイデルベルグレンシスとネアンデルタール人の分類上の区別は、海洋同位体段階8における30~243000年前のヨー ロッパでの化石ギャップに基づいてほとんど行われている。「しかし、シマ・デ・ロス・ウエソスの43万年前の骨は初期のネアンデルタール人または近縁のグ ループである可能性があり[24][138][139]、40万年前のアロエイラ3は過渡期である可能性がある。また、中期更新世において西ヨーロッパと アフリカの間で遺伝子流動があり、イタリアのセプラノやセルビアのシチェヴォ峡谷のような標本におけるネアンデルタール人の特徴を不明瞭にしていた可能性 もある[140]。 [イタリアのヴィソリアーノ遺跡とフォンタナ・ラヌッチョ遺跡からの歯列は、ネアンデルタール人の歯の特徴が中期更新世の約45万年から43万年前までに 進化していたことを示している[144][143]。 ネアンデルタール人と人間の分裂後のネアンデルタール人の進化に関して2つの主要な仮説がある:2段階と付加である。二相論はザール氷河のような単一の主 要な環境イベントがヨーロッパ人のハイデルベルゲンシスの体格と頑健性を急速に増加させ、頭部の長大化(第一段階)を引き起こし、それが頭蓋骨構造の他の 変化(第二段階)につながったと主張している[125] 。 66] 降着はネアンデルタール人が祖先のハイデルベルグエンシスから時間をかけてゆっくりと進化し、初期-前ネアンデルタール人(MIS 12、エルスター氷河期)、前ネアンデルタール感覚(MIS 11-9, Holstein間氷期)、初期ネアンデルタール(MIS 7-5, Saale 氷河期-エマ期)、古典ネアンデルタール感覚(MIS 4-3, Würm 氷河期)という4段階に分割したとする[136][136][137]。 ネアンデルタール人とヒトの分岐の年代は、これまでにも数多く提案されている。約25万年前という年代は、「H. helmei」を最後の共通祖先(LCA)とし、その分岐はレバロアの石器製作技術に関連しているとされる。約40万年前の年代は、「ハイデルベルクエン シス」をLCAとしている。60万年前の推定では、「H. rhodesiensis」をLCAとし、現代人系統とネアンデルタール人・ハイデルベルゲンシス人系統に分かれたとする[145] 80万年前の推定では「H. antecessor」をLCAとするが、このモデルの異なるバリエーションによって100万年前までさかのぼることになる[24][145] しかし2020年のH. antecessorエナメル質を用いた分析から、H. antecessorは、現代人とネアンデルタール人・ハイデルベルゲン人系統が関連していることが示唆されている。ネアンデルタール人とヒトの分岐時期 については、DNA研究によって538-315年[22] 553-321年[147] 565-503年[148] 654-475年[145] 690-550年[149] 765-550年[24] [89] 741-317年 [150] 800-52万年前など様々な結果が得られ、歯牙分析では80万年前以前という結論になっている[151]。 ネアンデルタール人とデニソワ人は現代人よりも近縁であり、ネアンデルタール人とデニソワ人の分裂は現代人との分裂後に起こったことを意味する[24] [89][133][152] 突然変異率を1×10-9または0. ネアンデルタール人とデニソワ人の分岐はそれぞれ236-19万年前と473-381万年前に発生した[89]。1年あたり5×10-10ヌクレオチドサ イトを用いると、616,000年前となる。後者の年代を用いると、ヨーロッパにホミニンが広がった時にはすでに分裂が起こっていた可能性が高く、ネアン デルタール人のユニークな特徴は60-50万年前までに進化し始めていた[133]。分裂の前に、アフリカからヨーロッパに移動したネアンデルタール人/ デニソワ人(または「ネアンデルソーバン」)は明らかにそこにすでに存在していた未確認「超archaic」人類種と混血しており、これらの超 archaicsはアフリカから約1.9myaに非常に早く移動していった末裔であった[153]。 |
| What does it mean to have
Neanderthal or Denisovan DNA? Several direct-to-consumer genetic testing companies report how much DNA a person has inherited from prehistoric humans, such as Neanderthals and Denisovans. This information is generally reported as a percentage that suggests how much DNA an individual has inherited from these ancestors. The percentage of Neanderthal DNA in modern humans is zero or close to zero in people from African populations, and is about 1 to 2 percent in people of European or Asian background. The percentage of Denisovan DNA is highest in the Melanesian population (4 to 6 percent), lower in other Southeast Asian and Pacific Islander populations, and very low or undetectable elsewhere in the world. Neanderthals were very early (archaic) humans who lived in Europe and Western Asia from about 400,000 years ago until they became extinct about 40,000 years ago. Denisovans are another population of early humans who lived in Asia and were distantly related to Neanderthals. (Much less is known about the Denisovans because scientists have uncovered fewer fossils of these ancient people.) The precise way that modern humans, Neanderthals, and Denisovans are related is still under study. However, research has shown that modern humans overlapped with Neanderthal and Denisovan populations for a period, and that they had children together (interbred). As a result, many people living today have a small amount of genetic material from these distant ancestors. Scientists have sequenced Neanderthal and Denisovan genomes from fossils discovered in Europe and Asia. This genetic information is helping researchers learn more about these early humans. Determining which areas of the genome are shared with archaic humans, and which areas are different, will also help researchers find out what differentiates modern humans from our closest extinct relatives. In addition to the percentage of Neanderthal or Denisovan DNA, direct to-consumer testing reports may include information about a few genetic variants inherited from these ancestors that influence specific traits. Some studies have suggested that certain genetic variations inherited from archaic humans may play roles in hair texture, height, sensitivity of the sense of smell, immune responses, adaptations to high altitude, and other characteristics in modern humans. These variations may also influence the risk of developing certain diseases. However, other studies have not found the same associations. The significance of Neanderthal or Denisovan genetic variants on disease risk is still an area of active study, and most direct-to-consumer test results currently do not include them. While knowing how much DNA a person has in common with his or her Neanderthal or Denisovan ancestors may be interesting, these data do not provide practical information about a person’s current health or chances of developing particular diseases. Having more or less DNA in common with archaic humans says nothing about how “evolved” a person is, nor does it give any indication of strength or intelligence. For now, knowing which specific genetic variants a person inherited from Neanderthal or Denisovan ancestors provides only limited information about a few physical traits. Scientific journal articles for further reading Meyer M, Kircher M, Gansauge MT, Li H, Racimo F, Mallick S, Schraiber JG, Jay F, Prüfer K, de Filippo C, Sudmant PH, Alkan C, Fu Q, Do R, Rohland N, Tandon A, Siebauer M, Green RE, Bryc K, Briggs AW, Stenzel U, Dabney J, Shendure J, Kitzman J, Hammer MF, Shunkov MV, Derevianko AP, Patterson N, Andrés AM, Eichler EE, Slatkin M, Reich D, Kelso J, Pääbo S. A high-coverage genome sequence from an archaic Denisovan individual. Science. 2012 Oct 12;338(6104):222-6. doi: 10.1126/science.1224344. Epub 2012 Aug 30. PubMed: 22936568; Free full-text article from PubMed Central: PMC3617501. Pääbo S. The diverse origins of the human gene pool. Nat Rev Genet. 2015 Jun;16(6):313-4. doi: 10.1038/nrg3954. PubMed: 25982166. Sankararaman S, Mallick S, Dannemann M, Prüfer K, Kelso J, Pääbo S, Patterson N, Reich D. The genomic landscape of Neanderthal ancestry in present-day humans. Nature. 2014 Mar 20;507(7492):354-7. doi: 10.1038/nature12961. Epub 2014 Jan 29. PubMed: 24476815. Free full-text article from PubMed Central: PMC4072735. Skov L, Coll Macià M, Sveinbjörnsson G, Mafessoni F, Lucotte EA, Einarsdóttir MS, Jonsson H, Halldorsson B, Gudbjartsson DF, Helgason A, Schierup MH, Stefansson K. The nature of Neanderthal introgression revealed by 27,566 Icelandic genomes. Nature. 2020 Jun;582(7810):78-83. doi: 10.1038/s41586-020-2225-9. Epub 2020 Apr 22. PubMed: 32494067. https://medlineplus.gov/genetics/understanding/dtcgenetictesting/neanderthaldna/ |
ネアンデルタール人やデニソワ人のDNAを持つということはどういうこ
とか? いくつかの消費者向け遺伝子検査会社は、ある人がネアンデルタール人やデニソワ人などの先史時代の人類からどの程度のDNAを受け継いでいるのかを報告し ている。この情報は通常、個人がこれらの祖先から受け継いだDNAの量を示すパーセンテージで報告される。現代人におけるネアンデルタール人のDNAの割 合は、アフリカ系住民ではゼロかそれに近く、ヨーロッパやアジア系住民では1~2%程度である。デニソワ人のDNAの割合はメラネシア人の集団で最も高く (4~6%)、他の東南アジアや太平洋諸島の集団では低く、世界の他の地域では非常に低いか検出されない程度である。 ネアンデルタール人は、約40万年前から約4万年前に絶滅するまで、ヨーロッパと西アジアに住んでいた非常に初期の(古代の)人類である。デニソワ人は、 アジアに住んでいた初期人類のもう一つの集団で、ネアンデルタール人と遠縁の関係にある(デニソワ人についてはあまり知られていない)。(デニソワ人につ いては、化石があまり発見されていないため、あまり知られていない。) 現代人、ネアンデルタール人、デニソワ人の正確な関係についてはまだ研究中である。しかし、現代人がネアンデルタール人やデニソワ人の集団と重なった時期 があり、一緒に子どもを産んだ(交配した)ことが研究で明らかになっている。その結果、現代に生きる多くの人々が、これらの遠い祖先の遺伝子を少量ずつ受 け継いでいる。 科学者たちは、ヨーロッパとアジアで発見された化石からネアンデルタール人とデニソワ人のゲノムを解読した。この遺伝子情報は、研究者がこれらの初期の人 類についてより詳しく知るために役立っている。ゲノムのどの部分が古代の人類と共通で、どの部分が異なるのかを明らかにすることは、研究者が現代人と最も 近い絶滅した親類を区別するポイントを見つけるのにも役立つ。 ネアンデルタール人やデニソワ人のDNAの割合に加えて、消費者向けの直接検査レポートには、これらの祖先から受け継いだ、特定の形質に影響を与えるいく つかの遺伝子変異についての情報が含まれていることがある。いくつかの研究では、古代の人類から受け継いだ特定の遺伝子変異が、現代人の髪質、身長、嗅覚 の敏感さ、免疫反応、高地への適応、その他の特徴に関与している可能性が示唆されている。また、これらの変異は、特定の病気の発症リスクにも影響している 可能性がある。しかし、他の研究では、同じような関連性は見つかっていない。ネアンデルタール人やデニソワ人の遺伝子変異が病気のリスクに与える影響につ いては、現在も活発に研究されている分野であり、現在、ほとんどの消費者向け直接検査結果には、これらの遺伝子変異が含まれていない。 ネアンデルタール人やデニソワ人の祖先と共通するDNAの量を知ることは興味深いことですが、これらのデータは、その人の現在の健康状態や特定の病気にか かる可能性についての実用的な情報を提供するものではありません。古代の人類と共通するDNAが多いか少ないかということは、その人がどれだけ「進化」し ているかを示すものではなく、体力や知性を示すものでもないのだ。今のところ、ネアンデルタール人やデニソワ人の祖先から受け継いだ特定の遺伝子変異を知 ることは、いくつかの身体的特徴についての限られた情報しか提供しない。  |
| ●ネアンデルタール人のDNAがCOVID-19の重症化リスクを高める
Nature ダイジェスト Vol. 18 No. 2 | doi : 10.1038/ndigest.2021.210244 原文:Nature (2020-11-26) | doi: 10.1038/d41586-020-02957-3 | Neanderthal DNA highlights complexity of COVID risk factors Yang Luo SARS-CoV-2感染に重度の反応を示す人々の中には、DNAのある部分をネアンデルタール人から受け継いでいる人がいることが、遺伝子解析によって 明らかにされた。ただし、ウイルスに対する反応様式の違いの全てがこの祖先によるわけではない。 COVID-19への取り組みでは、症状の程度に個人差がある理由を解明することが1つのカギになっている。2020年6月に発表された研究では、長さ5万ヌクレオチドのDNAセグメント(ヒトゲノムの 0.002%に相当)がCOVID-19の重症化や入院と強く関連することが明らかにされた1。そ のDNAセグメントがネアンデルタール人から受け継がれたものであることを、マックス・プランク進化人類学研究所(ドイツ・ライプチヒ)およびカロリンス カ研究所(スウェーデン・ストックホルム)のHugo Zebergと、マックス・プランク進化人類学研究所および沖縄科学技術大学院大学(沖縄県国頭郡恩納村)のSvante Pääbo2が、Nature 2020年11月26日号の610ページで明らかにした。このことにより、重症化しやすい人がいる理由の1 つが明らかになり、さらにはヒトの進化生物学に新たな知見がもたらされる。 ゲノム中で互いに近接しているDNA配列は一緒に受け継がれる(連鎖する)ことが多い。そのため、そうしたDNA の固まり(ハプロタイプ)には、密接に連鎖したバリアント(集団内の個人間で異なるDNA配列やヌクレオチド)が含まれる。例えば、上述のCOVID- 19重症化のリスクハプロタイプ1は、5万ヌクレオチドの配列の全体にわたって、98%以上の確率で一緒に受け継がれているバリアントを有している。この ような長いハプロタイプは、正の選択の結果なのかもしれない。ヒトの生存や生殖成功の都合に寄与したために、ゲノム中に維持された可能性があるのだ。さら にそれは、デニソワ人やネアンデルタール人などの旧人種との混血によってもたらされた可能性もある。 現代人のゲノムの約1~4%はそうした古い近縁種に由来している3。維 持されている旧人遺伝子の多くは現代人にとって有害であり、生殖能力の低下や高い疾患リスクと関連付けられている4。しかし、有 益なものもわずかにある。例えば、EPAS1と呼ばれる遺伝子のデニソワ人様のバリアントは、現代のチベット人が極端な高地での生活に対処するのに役立っ ており5、ネアンデルタール人の遺伝子には、疼痛感受性を高めているもの6やウイルスから身を守るのに役立っているもの7もある。 COVID-19重症化のリスクハプロタイプが旧人からもたらされたのかどうかを調べるため、ZebergとPääboは、この領域を世界各地の旧人ゲノ ムのオンラインデータベースと照合した。その結果、この領域は、約5万年前に現在の クロアチアに住んでいたあるネアンデルタール人の当該ゲノム領域に近い一方、既知のどのデニソワ人ゲノムとも近縁ではないことが明らかになった。 次にZebergとPääboは、ネアンデルタール人由来のこのハプロタイプが現代人集団内にどれだけ広がっているかを調べた。すると、東 アジア人とアフリカ人にはほとんどあるいは全く認められず、ラテンアメリカ人と欧州人では低い頻度(それぞれ4%および8%)で維持されていることが分 かった。対照的に、南アジア系の人々にはこのハプロタイプが30%の頻度で存在し、バングラデシュ系では37%にも上った(図1)。 ++++++++++++++++ 図1 COVID-19の重症化に関連する遺伝的リスク因子の不均一な世界的分布 ZebergとPääbo2は、COVID-19の重症化や入院と関連する長いDNA配列がネアンデルタール人に由来していることを明らかにした。その配 列は現代人集団の間で不均一に分布している。この地図は、世界各地のさまざまな集団にそのリスク因子がどれだけの頻度で見られるかを示している。各集団の 塩基配列解読データは1000ゲノムプロジェクトによって収集された10(参考文献2のFig. 3を改変)。 ++++++++++++++++ これによりZebergとPääboは、特定の集団ではネアンデルタール人由来のこ のハプロタイプがCOVID-19重症化リスクの大きな一因になっているのではないかと考えた。その仮説は英国国家統計局の病院データ8によって支持され る。同国のバングラデシュ系の人々のCOVID-19による死亡率が、一般集団の2倍に達することが示されているのだ(ただし、こうした統計にはもちろん 他のリスク因子が寄与していることも考えられる)。 なぜ、一部の集団ではこのハプロタイプが維持されてきたのだろうか? ZebergとPääboは、他の古い病原体から身を守っていたために、世界各地の特定集団で正の選択を受けた9のではないかと考えている。し かし、SARS-CoV-2感染では、そうした旧人の遺伝子が引き起こす防御免疫反応が過度に攻撃的となり、COVID-19が重症化する人々に見られる 命に関わりかねない免疫反応を生じているのかもしれない。結果として、過去には生存に有益だったのではないかと考えられるハプロタイプが、今は逆効果に なっている可能性がある。 このリスクハプロタイプと臨床転帰には相関はあるが、個人のCOVID-19重症化リスクは遺伝学だけで決まるわけではない。ヒトの遺伝子とその起源は、 COVID-19(および他の感染症)の発症と進行に間違いなく影響を与えるが、疾患の転帰には環境因子も大きな役目を果たす。 例えば、アフリカ系の人々にはネアンデルタール人由来のこのリスクハプロタイプがほぼ全く存在しないが、地理的、社会経済的因子に関する調整を行っても、 彼らは他の人種的背景を持つ人々よりもCOVID-19の死亡率が高い(go.nature.com/3jcxezx[Demographicsのタブ] およびgo.nature.com/2h4qfquなどを参照)。COVID-19の死亡リスクに関して、社会的不平等とその影響は、ネアンデルタール人 由来のDNAよりも大きな要因となっているように思われる。 我々の祖先の遺伝的遺産が現在のパンデミックで責を負っているかもしれ ない、と考えることには興味をそそられる。しかし、受け継がれたDNAがウイルスに対する体の反応に与える根源的な影響は明らかにされていない。COVID -19宿主遺伝学イニシアチブ(www.covid19hg.org) が進めているような、多様な集団に属するさらに多くの個人を解析することによって遺伝学とCOVID-19の病態との関連を研究する現下の世界的取り組み は、原因解明をさらに進めるのに役立つと考えられる。COVID-19の病態に関与する遺伝子は受け継がれているかもしれない。しかし重要なのは、我々に 制御できることは社会的因子と行動(ソーシャルディスタンスの確保やマスクの着用など)であり、それによって感染リスクが効果的に抑制可能であると認める ことだ。 (翻訳:小林盛方) The Severe Covid-19 GWAS Group. N. Engl. J. Med. 383, 1522–1534 (2020). Zeberg, H. & Pääbo, S. Nature 587, 610–612 (2020). Green, R. E. et al. Science 328, 710–722 (2010). Sankararaman, S. et al. Nature 507, 354–357 (2014). Huerta-Sánchez, E. et al. Nature 512, 194–197 (2014). Zeberg, H. et al. Curr. Biol. 30, 3465–3469 (2020). Enard, D. & Petrov, D. A. Cell 175, 360–371 (2018). Public Health England. Disparities in the Risk and Outcomes of COVID-19 (PHE Publs, 2020). Browning, S. R., Browning, B. L., Zhou, Y., Tucci, S. & Akey, J. M. Cell 173, 53–61 (2018). The 1000 Genomes Project Consortium. Nature 526, 68–74 (2015). https://go.nature.com/3VFhj9G |
|
| Interbreeding Main article: Archaic human admixture with modern humans Interbreeding with modern humans Further information: Neanderthal genetics 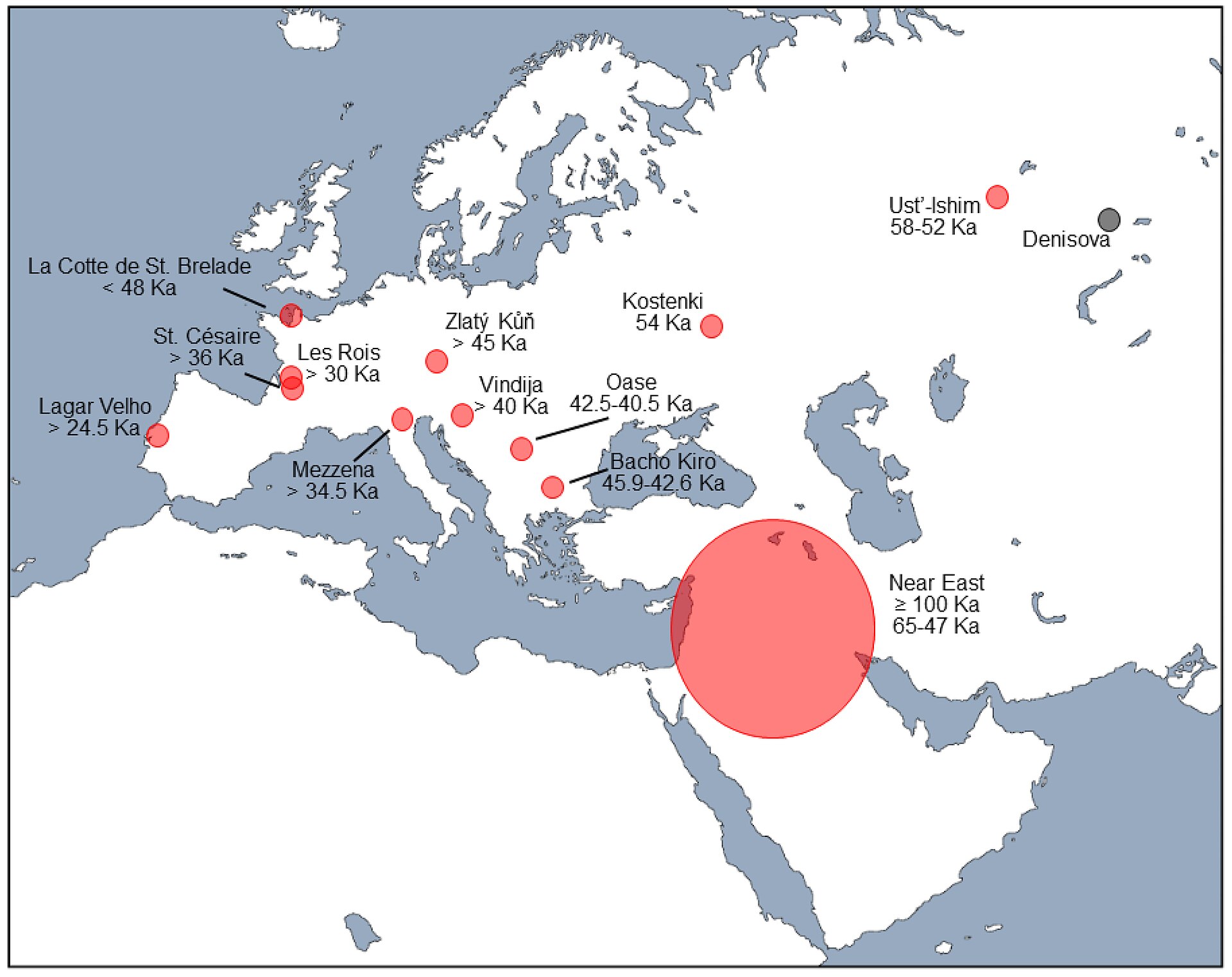 Map of western Eurasia showing areas and estimated dates of possible Neandertal–modern human hybridisation (in red) based on fossil samples from indicated sites[400] The first Neanderthal genome sequence was published in 2010, and strongly indicated interbreeding between Neanderthals and early modern humans.[87][401][402][403] The genomes of all studied modern populations contain Neanderthal DNA.[87][89][404][405][406] Various estimates exist for the proportion, such as 1–4%[87] or 3.4–7.9% in modern Eurasians,[407] or 1.8–2.4% in modern Europeans and 2.3–2.6% in modern East Asians.[408] Pre-agricultural Europeans appear to have had similar, or slightly higher,[406] percentages to modern East Asians, and the numbers may have decreased in the former due to dilution with a group of people which had split off before Neanderthal introgression.[100] Typically, studies have reported finding no significant levels of Neanderthal DNA in Sub-Saharan Africans, but a 2020 study detected 0.3-0.5% in the genomes of five African sample populations, likely the result of Eurasians back-migrating and interbreeding with Africans, as well as human-to-neanderthal gene flow from dispersals of Homo sapiens preceding the larger Out-of-Africa migration, and also showed more equal Neanderthal DNA percentages for European and Asian populations.[406] Such low percentages of Neanderthal DNA in all present day populations indicate infrequent past interbreeding,[409] unless interbreeding was more common with a different population of modern humans which did not contribute to the present day gene pool.[100] Of the inherited Neanderthal genome, 25% in modern Europeans and 32% in modern East Asians may be related to viral immunity.[410] In all, approximately 20% of the Neanderthal genome appears to have survived in the modern human gene pool.[94]  A dark-skinned man with black, shiny hair going down to his shoulders, a slight moustache, a goatee, brown eyes, weak eyebrows, wearing a tailored shirt and holding a long spear to support himself Reconstruction of the upper Palaeolithic human Oase 2 with around 7.3% Neanderthal DNA (from an ancestor 4–6 generations back)[411] However, due to their small population and resulting reduced effectivity of natural selection, Neanderthals accumulated several weakly harmful mutations, which were introduced to and slowly selected out of the much larger modern human population; the initial hybridised population may have experienced up to a 94% reduction in fitness compared to contemporary humans. By this measure, Neanderthals may have substantially increased in fitness.[82] A 2017 study focusing on archaic genes in Turkey found associations with coeliac disease, malaria severity and Costello syndrome.[412] Nonetheless, some genes may have helped modern East Asians adapt to the environment; the putatively Neanderthal Val92Met variant of the MC1R gene, which may be weakly associated with red hair and UV radiation sensitivity,[413] is primarily found in East Asian, rather than European, individuals.[414] Some genes related to the immune system appear to have been affected by introgression, which may have aided migration,[96] such as OAS1,[415] STAT2,[416] TLR6, TLR1, TLR10,[417] and several related to immune response.[95][f] In addition, Neanderthal genes have also been implicated in the structure and function of the brain,[g] keratin filaments, sugar metabolism, muscle contraction, body fat distribution, enamel thickness and oocyte meiosis.[99] Nonetheless, a large portion of surviving introgression appears to be non-coding ("junk") DNA with few biological functions.[100] There is considerably less Neanderthal ancestry on the X-chromosome as compared to the autosomal chromosomes. This has led to suggestions that admixture with modern humans was sex biased, and primarily the result of mating between modern human females and Neanderthal males. Other authors have suggested that this may be due to negative selection against Neandethal alleles, however these two proposals are not mutually exclusive.[141] A 2023 study confirmed that the low level of Neanderthal ancestry on the X-chromosomes is best explained by sex bias in the admixture events, and these authors also found evidence for negative selection on archaic genes.[419] Neanderthal mtDNA (which is passed on from mother to child) is absent in modern humans.[136][155][420] This is evidence that interbreeding occurred mainly between Neanderthal males and modern human females.[421] According to Svante Pääbo, it is not clear that modern humans were socially dominant over Neanderthals, which may explain why the interbreeding occurred primarily between Neanderthal males and modern human females.[422] Furthermore, even if Neanderthal women and modern human males had interbred, Neanderthal mtDNA lineages may have gone extinct if women who carried them only gave birth to sons.[422] Due to the lack of Neanderthal-derived Y-chromosomes in modern humans (which is passed on from father to son), it has also been suggested that the hybrids that contributed ancestry to modern populations were predominantly females, or the Neanderthal Y-chromosome was not compatible with H. sapiens and became extinct.[100][423] According to linkage disequilibrium mapping, the last Neanderthal gene flow into the modern human genome occurred 86–37,000 years ago, but most likely 65–47,000 years ago.[424] It is thought that Neanderthal genes which contributed to the present day human genome stemmed from interbreeding in the Near East rather than the entirety of Europe. However, interbreeding still occurred without contributing to the modern genome.[100] The approximately 40,000-year-old modern human Oase 2 was found, in 2015, to have had 6–9% (point estimate 7.3%) Neanderthal DNA, indicating a Neanderthal ancestor up to four to six generations earlier, but this hybrid population does not appear to have made a substantial contribution to the genomes of later Europeans.[411] In 2016, the DNA of Neanderthals from Denisova Cave revealed evidence of interbreeding 100,000 years ago, and interbreeding with an earlier dispersal of H. sapiens may have occurred as early as 120,000 years ago in places such as the Levant.[91] The earliest H. sapiens remains outside of Africa occur at Misliya Cave 194–177,000 years ago, and Skhul and Qafzeh 120–90,000 years ago.[425] The Qafzeh humans lived at approximately the same time as the Neanderthals from the nearby Tabun Cave.[426] The Neanderthals of the German Hohlenstein-Stadel have deeply divergent mtDNA compared to more recent Neanderthals, possibly due to introgression of human mtDNA between 316,000 and 219,000 years ago, or simply because they were genetically isolated.[90] Whatever the case, these first interbreeding events have not left any trace in modern human genomes.[427] Genetic evidence suggests that following their split from Denisovans, Neanderthals experienced gene flow (around 3% of their genome) from the lineage leading to modern humans prior to the expansion of modern humans outside of Africa during the Last Glacial Period, with this interbreeding suggested to have taken place around 200-300,000 years ago.[141] Detractors of the interbreeding model argue that the genetic similarity is only a remnant of a common ancestor instead of interbreeding,[428] although this is unlikely as it fails to explain why sub-Saharan Africans do not have Neanderthal DNA.[402] Interbreeding with Denisovans 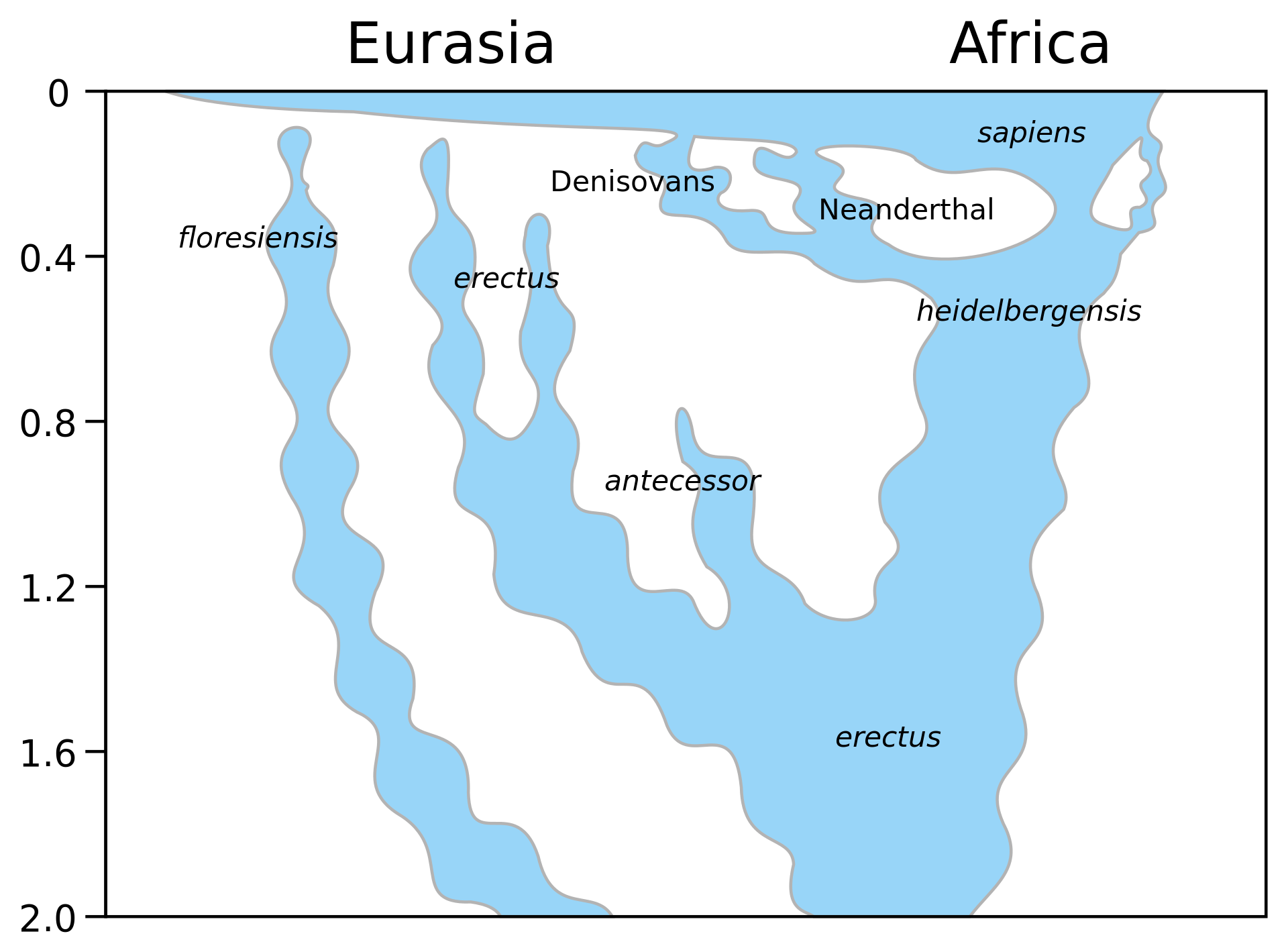 Chris Stringer's Homo family tree. The horizontal axis represents geographic location, and the vertical time in millions of years ago.[h] Although nDNA confirms that Neanderthals and Denisovans are more closely related to each other than they are to modern humans, Neanderthals and modern humans share a more recent maternally-transmitted mtDNA common ancestor, possibly due to interbreeding between Denisovans and some unknown human species. The 400,000-year-old Neanderthal-like humans from Sima de los Huesos in northern Spain, looking at mtDNA, are more closely related to Denisovans than Neanderthals. Several Neanderthal-like fossils in Eurasia from a similar time period are often grouped into H. heidelbergensis, of which some may be relict populations of earlier humans, which could have interbred with Denisovans.[430] This is also used to explain an approximately 124,000-year-old German Neanderthal specimen with mtDNA that diverged from other Neanderthals (except for Sima de los Huesos) about 270,000 years ago, while its genomic DNA indicated divergence less than 150,000 years ago.[90] Sequencing of the genome of a Denisovan from Denisova Cave has shown that 17% of its genome derives from Neanderthals.[93] This Neanderthal DNA more closely resembled that of a 120,000-year-old Neanderthal bone from the same cave than that of Neanderthals from Vindija Cave, Croatia, or Mezmaiskaya Cave in the Caucasus, suggesting that interbreeding was local.[92] For the 90,000-year-old Denisova 11, it was found that her father was a Denisovan related to more recent inhabitants of the region, and her mother a Neanderthal related to more recent European Neanderthals at Vindija Cave, Croatia. Given how few Denisovan bones are known, the discovery of a first-generation hybrid indicates interbreeding was very common between these species, and Neanderthal migration across Eurasia likely occurred sometime after 120,000 years ago.[431] |
異種交配 主な記事 古人類と現生人類の交配 現代人との交雑 さらなる情報 ネアンデルタール人の遺伝学 ネアンデルタール人と現生人類が交雑した可能性のある地域と推定年代を示すユーラシア大陸西部の地図(赤)[400]。 2010年に最初のネアンデルタール人のゲノム配列が発表され、ネアンデルタール人と初期の現代人との交雑が強く示唆された[87][401][402] [403]。 [87][89][404][405][406]現代ユーラシア人においては1-4%[87]、3.4-7.9%[407]、現代ヨーロッパ人においては 1.8-2.4%、現代東アジア人においては2.3-2.6%など、その割合については様々な推定が存在する。 [408]農業以前のヨーロッパ人は、現代の東アジア人と同程度か、わずかに高い割合[406]を有していたようであり、ネアンデルタール人の導入以前に 分かれた人々のグループとの希釈により、前者ではその数が減少した可能性がある[100]。一般的に、サハラ以南のアフリカ人においては、有意なレベルの ネアンデルタール人DNAは検出されなかったと報告されているが、2020年の研究では、5人のゲノムの0.3-0.5%が検出された。 これはおそらくユーラシア人がアフリカ人と逆移行して交雑した結果であり、またアフリカ外への大移動に先立つホモ・サピエンスの分散からのヒトからネアン デルタール人への遺伝子流動の結果でもある。 [406] 現在のすべての集団におけるネアンデルタール人DNAのこのような低い割合は、現在の遺伝子プールに寄与していない現生人類の別の集団との交雑がより一般 的でない限り、過去の交雑の頻度が低かったことを示している[409]。 黒く輝く髪を肩まで伸ばし、わずかに口ひげを生やし、あごひげを生やし、茶色の目、弱々しい眉毛を持ち、仕立てたシャツを着て、長い槍を持って体を支えて いる。 約7.3%のネアンデルタール人のDNA(4~6世代前の祖先から)を持つ旧石器時代後期の人類オアーゼ2の復元[411]。 しかし、ネアンデルタール人は、その小さな集団とその結果としての自然淘汰の有効性の低下により、いくつかの弱く有害な突然変異を蓄積し、それがはるかに 大きな現代人の集団に導入され、ゆっくりと淘汰されていった。この尺度からすると、ネアンデルタール人の体力は実質的に向上していた可能性がある [82]。2017年にトルコで行われた古代の遺伝子に焦点を当てた研究では、セリアック病、マラリアの重症度、コステロ症候群との関連が発見された [412]。それにもかかわらず、いくつかの遺伝子は現代の東アジア人の環境への適応を助けた可能性がある。赤毛や紫外線感受性[413]と弱く関連して いる可能性があるMC1R遺伝子のネアンデルタール人とされるVal92Met変異体は、主にヨーロッパ人ではなく東アジア人で発見されている。 [OAS1, [415] STAT2, [416] TLR6, TLR1, TLR10, [417]および免疫反応に関連するいくつかの遺伝子など[96]。 [95][f]さらに、ネアンデルタール人の遺伝子は、脳の構造と機能、[g]ケラチンフィラメント、糖代謝、筋肉収縮、体脂肪分布、エナメル質の厚さ、 卵子の減数分裂にも関与している。 常染色体に比べてX染色体にはネアンデルタール人の祖先がかなり少ない。このため、現生人類との混血は性に偏りがあり、主に現生人類の女性とネアンデル タール人の男性との交配の結果であるという指摘がある。他の著者は、これはネアンデルタール人の対立遺伝子に対する負の選択によるものではないかと示唆し ているが、これら2つの提案は相互に排他的なものではない。2023年の研究では、X染色体上のネアンデルタール人の祖先のレベルの低さは、混血事象にお ける性の偏りによって最もよく説明できることが確認され、これらの著者は古代の遺伝子に対する負の選択の証拠も発見した[419]。 ネアンデルタール人のmtDNA(母から子へ受け継がれる)は現生人類には存在しない[136][155][420]。これは主にネアンデルタール人の男 性と現生人類の女性の間で交雑が起こったという証拠である。 [421] スヴァンテ・ペーボによれば、現生人類がネアンデルタール人よりも社会的に支配的であったことは明らかではなく、それがネアンデルタール人の男性と現生人 類の女性の間で主に交雑が起こった理由を説明しているのかもしれない[422]。さらに、ネアンデルタール人の女性と現生人類の男性が交雑していたとして も、それを持つ女性が息子しか産まなければ、ネアンデルタール人のmtDNA系統は絶滅していたかもしれない[422]。 現生人類にはネアンデルタール人由来のY染色体が存在しない(これは父から子へと受け継がれる)ことから、現生人類集団に祖先をもたらした雑種は主に女性 であった、あるいはネアンデルタール人のY染色体はH.サピエンスと適合せず絶滅したとも考えられている[100][423]。 連鎖不平衡マッピングによると、ネアンデルタール人の遺伝子が現生人類のゲノムに流入したのは86-37,000年前であるが、最も可能性が高いのは65 -47,000年前である[424]。100]約4万年前の現生人類Oase 2は、2015年にネアンデルタール人のDNAを6-9%(推定値7.3%)持っていたことが発見され、これはネアンデルタール人の祖先が4-6世代前ま でいたことを示しているが、このハイブリッド集団は後のヨーロッパ人のゲノムに実質的な貢献はしていないようである。 [2016年、デニソワ洞窟から採取されたネアンデルタール人のDNAから10万年前の交雑の証拠が明らかになった。 [アフリカ以外で最古のH.サピエンスの遺跡は、194-177,000年前のミスリヤ洞窟と120-90,000年前のスクルとカフゼで発見されてい る。 [426]ドイツのホーレンシュタイン=シュターデルのネアンデルタール人は、より最近のネアンデルタール人と比べてmtDNAが深く乖離しているが、こ れはおそらく316,000年から219,000年前の間にヒトのmtDNAが導入されたためか、あるいは単に遺伝的に隔離されていたためである。 遺伝学的証拠は、デニソワ人からの分裂後、ネアンデルタール人が最終氷期においてアフリカの外に現生人類が拡大する前に、現生人類につながる系統からの遺 伝子流入(ゲノムの約3%)を経験したことを示唆しており、この交雑は約20-30万年前に起こったと示唆されている[141]。 異種交配モデルの否定派は、遺伝的な類似性は異種交配ではなく共通の祖先の名残に過ぎないと主張しているが[428]、サハラ以南のアフリカ人がネアンデ ルタール人のDNAを持っていない理由を説明できないので、これはあり得ないことである[402]。 デニソワ人との交雑 クリス・ストリンガーのホモ家系図。横軸は地理的位置、縦軸は数百万年前の時間を表す[h]。 nDNAはネアンデルタール人とデニソワ人が現生人類よりも近縁であることを裏付けているが、ネアンデルタール人と現生人類は、おそらくデニソワ人と未知 の人類種との交雑に起因する、より新しい母方伝達型mtDNA共通祖先を共有している。スペイン北部のシマ・デ・ロス・ウエソスから発見された40万年前 のネアンデルタール人類似の人類は、mtDNAで見ると、ネアンデルタール人よりもデニソワ人に近縁である。このことは、約12万4千年前のドイツのネア ンデルタール人標本のmtDNAが他のネアンデルタール人(シマ・デ・ロス・ウエソスを除く)とは約27万年前に分岐しているのに対して、そのゲノム DNAは15万年未満前に分岐したことを示していることの説明にも使われている[90]。 デニソワ洞窟から採取されたデニソワ人のゲノムの塩基配列を決定したところ、そのゲノムの17%がネアンデルタール人に由来することが示された[93]。 このネアンデルタール人のDNAは、クロアチアのヴィンディヤ洞窟やコーカサスのメズマイスカヤ洞窟から採取されたネアンデルタール人のDNAよりも、同 じ洞窟から採取された12万年前のネアンデルタール人の骨のDNAに酷似しており、交雑が局地的であったことを示唆している[92]。 90,000年前のデニソワ11世については、父親がこの地域の最近の住民に関係するデニソワ人であり、母親がクロアチアのヴィンディヤ洞窟の最近のヨー ロッパのネアンデルタール人に関係するネアンデルタール人であることが判明した。デニソワ人の骨がほとんど知られていないことを考えると、第一世代の雑種 が発見されたことは、これらの種の間で交雑が非常に一般的であったことを示しており、ユーラシア大陸を横断するネアンデルタール人の移動は、おそらく12 万年前以降に起こったと考えられる[431]。 |
| Extinction Main article: Neanderthal extinction Transition  Black and white satellite image of the Iberian Peninsula, but the Ebro River valley at the Spain/France border uses red to blue colours to indicate topography and elevation Map emphasising the Ebro River in northern Spain The extinction of Neanderthals was part of the broader Late Pleistocene megafaunal extinction event.[432] Whatever the cause of their extinction, Neanderthals were replaced by modern humans, indicated by near full replacement of Middle Palaeolithic Mousterian stone technology with modern human Upper Palaeolithic Aurignacian stone technology across Europe (the Middle-to-Upper Palaeolithic Transition) from 41,000 to 39,000 years ago.[8][9][11][433] By between 44,200 to 40,600 BP, Neanderthals vanished from northwestern Europe.[434] However, it is postulated that Iberian Neanderthals persisted until about 35,000 years ago, as indicated by the date range of transitional lithic assemblages—Châtelperronian, Uluzzian, Protoaurignacian and Early Aurignacian. The latter two are attributed to modern humans, but the former two have unconfirmed authorship, potentially products of Neanderthal/modern human cohabitation and cultural transmission. Further, the appearance of the Aurignacian south of the Ebro River has been dated to roughly 37,500 years ago, which has prompted the "Ebro Frontier" hypothesis which states that the river presented a geographic barrier preventing modern human immigration, and thus prolonging Neanderthal persistence.[435][436] However, the dating of the Iberian Transition is debated, with a contested timing of 43,000–40,800 years ago at Cueva Bajondillo, Spain.[437][438][439][440] The Châtelperronian appears in northeastern Iberia about 42,500–41,600 years ago.[435] Some Neanderthals in Gibraltar were dated to much later than this—such as Zafarraya (30,000 years ago)[441] and Gorham's Cave (28,000 years ago)[442]—which may be inaccurate as they were based on ambiguous artefacts instead of direct dating.[11] A claim of Neanderthals surviving in a polar refuge in the Ural Mountains[169] is loosely supported by Mousterian stone tools dating to 34,000 years ago from the northern Siberian Byzovaya site at a time when modern humans may not yet have colonised the northern reaches of Europe;[171] however, modern human remains are known from the nearby Mamontovaya Kurya site dating to 40,000 years ago.[443] Indirect dating of Neanderthals remains from Mezmaiskaya Cave reported a date of about 30,000 years ago, but direct dating instead yielded 39,700 ±1,100 years ago, more in line with trends exhibited in the rest of Europe.[10]  Bohunician scrapers in the Moravian Museum, Czech Republic The earliest indication of Upper Palaeolithic modern human immigration into Europe is the Balkan Bohunician industry beginning 48,000 years ago, likely deriving from the Levantine Emiran industry,[355] and the earliest bones in Europe date to roughly 45–43,000 years ago in Bulgaria,[444] Italy,[445] and Britain.[446] This wave of modern humans replaced Neanderthals.[8] However, Neanderthals and H. sapiens have a much longer contact history. DNA evidence indicates H. sapiens contact with Neanderthals and admixture as early as 120–100,000 years ago. A 2019 reanalysis of 210,000-year-old skull fragments from the Greek Apidima Cave assumed to have belonged to a Neanderthal concluded that they belonged to a modern human, and a Neanderthal skull dating to 170,000 years ago from the cave indicates H. sapiens were replaced by Neanderthals until returning about 40,000 years ago.[447] This identification was refuted by a 2020 study.[448] Archaeological evidence suggests that Neanderthals displaced modern humans in the Near East around 100,000 years ago until about 60–50,000 years ago.[100] Cause Modern humans 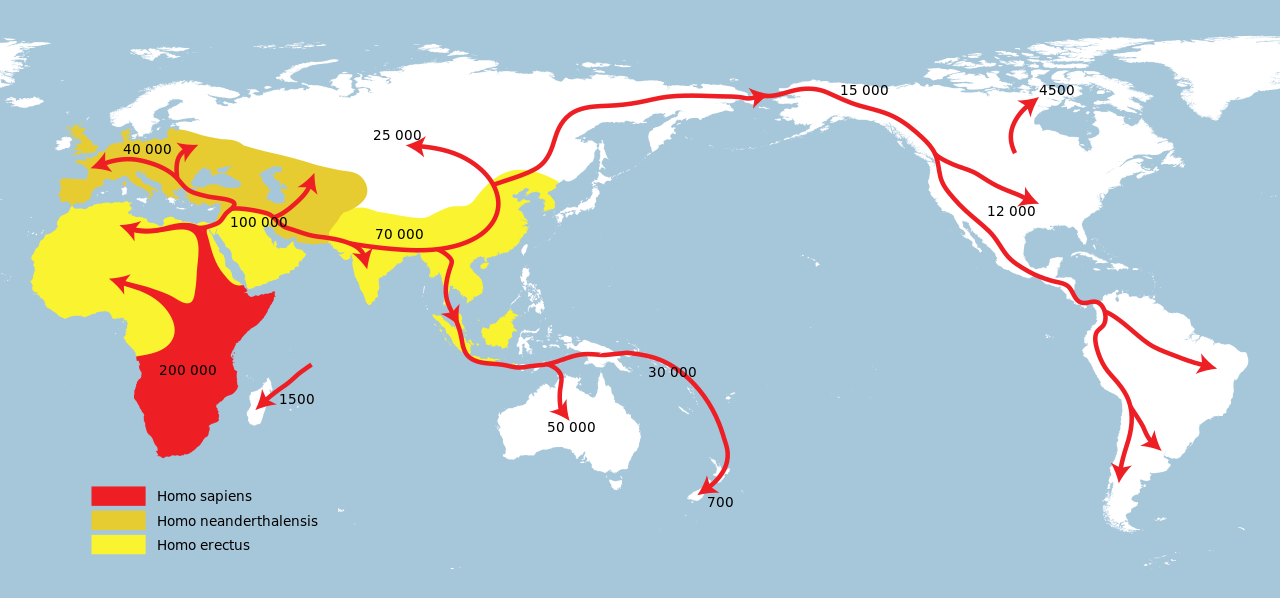 Successive dispersals of Homo erectus (yellow), Neanderthals (ochre) and modern humans (red). Historically, modern human technology was viewed as vastly superior to that of Neanderthals, with more efficient weaponry and subsistence strategies, and Neanderthals simply went extinct because they could not compete.[19] The discovery of Neanderthal/modern human introgression has caused the resurgence of the multiregional hypothesis, wherein the present day genetic makeup of all humans is the result of complex genetic contact among several different populations of humans dispersed across the world. By this model, Neanderthals and other recent archaic humans were simply assimilated into the modern human genome – that is, they were effectively bred out into extinction.[19] Modern humans coexisted with Neanderthals in Europe for around 3,000 to 5,000 years.[449] Climate change Their ultimate extinction coincides with Heinrich event 4, a period of intense seasonality; later Heinrich events are also associated with massive cultural turnovers when European human populations collapsed.[20][21] This climate change may have depopulated several regions of Neanderthals, like previous cold spikes, but these areas were instead repopulated by immigrating humans, leading to Neanderthal extinction.[450] In southern Iberia, there is evidence that Neanderthal populations declined during H4 and the associated proliferation of Artemisia-dominated desert-steppes.[451] 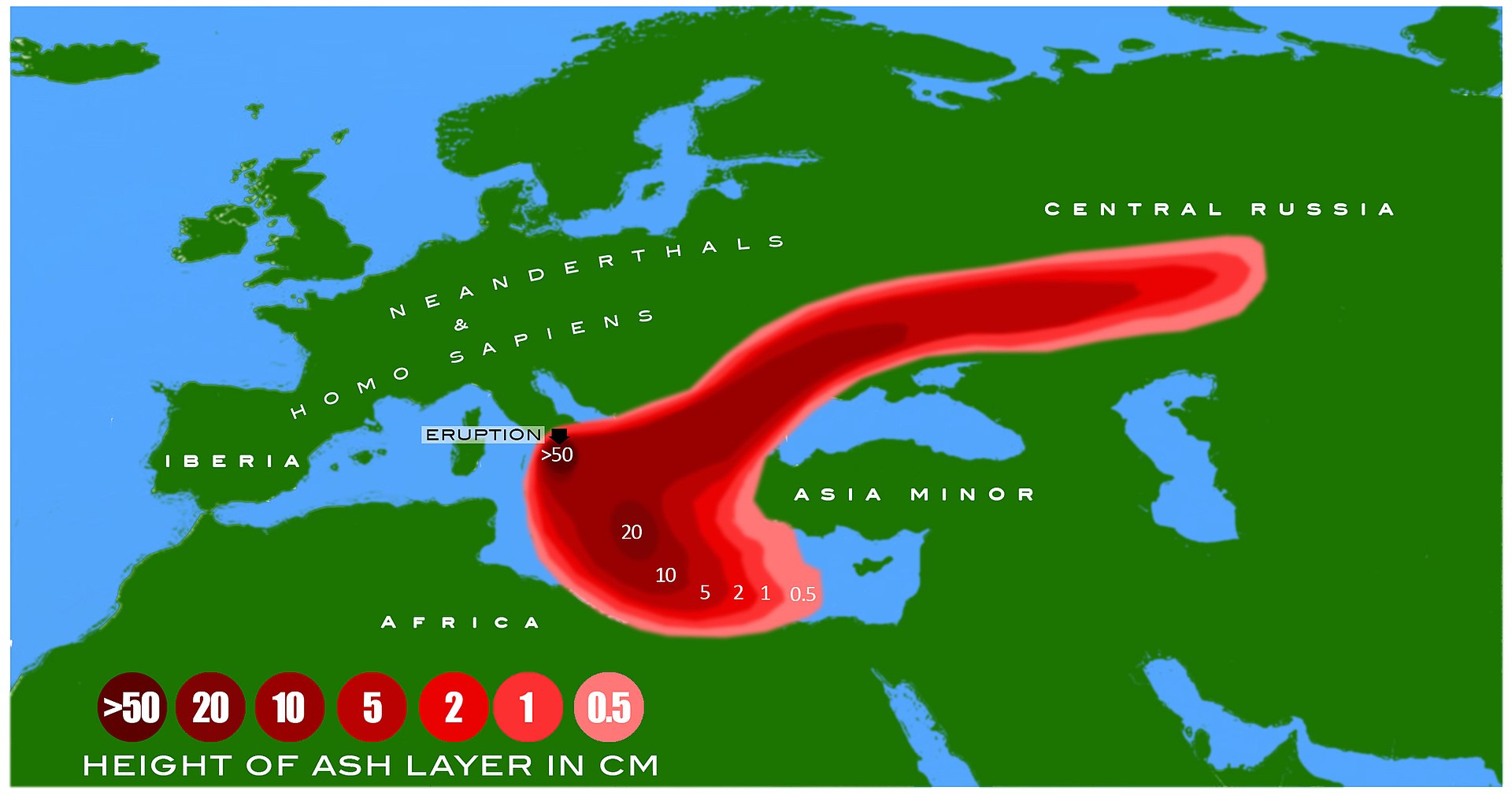 Map of Europe showing a red splotch over the Italian Peninsula stretching out to the east over Central Russia and over the Aegean and Ionic Seas, 50 cm of ash centring, decreasing to 10 cm beyond, and steadily decreasing to 0 Dispersal of deposits during the Campanian Ignimbrite Eruption around 40,000 years ago. It has also been proposed that climate change was the primary driver, as their low population left them vulnerable to any environmental change, with even a small drop in survival or fertility rates possibly quickly leading to their extinction.[452] However, Neanderthals and their ancestors had survived through several glacial periods over their hundreds of thousands of years of European habitation.[279] It is also proposed that around 40,000 years ago, when Neanderthal populations may have already been dwindling from other factors, the Campanian Ignimbrite Eruption in Italy could have led to their final demise, as it produced 2–4 °C (3.6–7.2 °F) cooling for a year and acid rain for several more years.[22][453] Disease Modern humans may have introduced African diseases to Neanderthals, contributing to their extinction. A lack of immunity, compounded by an already low population, was potentially devastating to the Neanderthal population, and low genetic diversity could have also rendered fewer Neanderthals naturally immune to these new diseases ("differential pathogen resistance" hypothesis). However, compared to modern humans, Neanderthals had a similar or higher genetic diversity for 12 major histocompatibility complex (MHC) genes associated with the adaptive immune system, casting doubt on this model.[24] Low population and inbreeding depression may have caused maladaptive birth defects, which could have contributed to their decline (mutational meltdown).[243] In late-20th-century New Guinea, due to cannibalistic funerary practices, the Fore people were decimated by transmissible spongiform encephalopathies, specifically kuru, a highly virulent disease spread by ingestion of prions found in brain tissue. However, individuals with the 129 variant of the PRNP gene were naturally immune to the prions. Studying this gene led to the discovery that the 129 variant was widespread among all modern humans, which could indicate widespread cannibalism at some point in human prehistory. Because Neanderthals are known to have practised cannibalism to an extent and to have co-existed with modern humans, British palaeoanthropologist Simon Underdown speculated that modern humans transmitted a kuru-like spongiform disease to Neanderthals, and, because the 129 variant appears to have been absent in Neanderthals, it quickly killed them off.[23][454] |
絶滅 主な記事 ネアンデルタール人の絶滅 変遷 イベリア半島の白黒衛星写真だが、スペインとフランスの国境にあるエブロ川流域は、地形と標高を示すために赤から青への色が使われている。 スペイン北部のエブロ川を強調した地図 ネアンデルタール人の絶滅は、より広範な更新世後期の巨大動物絶滅現象の一部であった[432]。その絶滅の原因が何であれ、ネアンデルタール人が現生人 類に取って代わられたことは、41,000年前から39,000年前にかけて、ヨーロッパ全域で、中期旧石器時代のモウステリアン石器技術が現生人類の上 部旧石器時代のアウリニャシアン石器技術にほぼ完全に置き換わったこと(中期から上部旧石器時代への移行)によって示されている。 [しかし、イベリア半島のネアンデルタール人は約35,000年前まで生存していたと推測されており、そのことは、シャテルペロニアン、ウルーズィアン、 プロトオウリニャシアン、初期アウリニャシアンといった移行期の石器群の年代範囲によって示されている。後者2つは現生人類によるものとされているが、前 者2つは作者が未確認であり、ネアンデルタール人と現生人類の同居と文化伝播の産物である可能性がある。さらに、エブロ川以南にアウリニャキア人が出現し たのはおよそ37,500年前とされており、エブロ川が現代人の移住を阻む地理的障壁となり、ネアンデルタール人の存続を長引かせたとする「エブロ・フロ ンティア」仮説が提唱されている[435][436]。 [437][438][439][440]。イベリア半島北東部ではシャテルペロニアンが約42,500-41,600年前に出現している[435]。 ジブラルタルにいたネアンデルタール人の中には、ザファラヤ(30,000年前)[441]やゴーラムの洞窟(28,000年前)[442]など、これよ りもかなり後の年代とされたものもあるが、これは直接的な年代測定ではなく曖昧な遺物に基づいているため、不正確である可能性がある。 [11] ネアンデルタール人がウラル山脈の極地避難所に生存しているという主張[169]は、現生人類がまだヨーロッパ北部を植民地化していなかったかもしれない 時期にシベリア北部のビゾヴァヤ遺跡から出土した34,000年前のモウステリアン石器によって緩やかに裏付けられている[171]。 [443]メズマイスカヤ洞窟から出土したネアンデルタール人の遺体の間接年代測定では約3万年前と報告されたが、直接年代測定では39,700± 1,100年前となり、ヨーロッパの他の地域で見られる傾向と一致した[10]。 チェコ共和国、モラヴィア博物館所蔵のボヒュニキア時代のスクレイパー ヨーロッパへの後期旧石器時代の現生人類の移住の最も早い兆候は、48,000年前に始まったバルカン半島のボフニキア産業であり、おそらくレヴァントの エミラン産業から派生したものであり[355]、ヨーロッパにおける最古の骨は、ブルガリア、[444]イタリア、[445]イギリスにおけるおよそ 45~43,000年前のものである[446]。DNAの証拠は、H.サピエンスがネアンデルタール人と接触し、混血したことを12-10万年前にも示し ている。2019年にギリシャのアピディマ洞窟でネアンデルタール人のものとされた21万年前の頭蓋骨片を再分析したところ、それは現生人類のものであっ たと結論づけられた。サピエンスは約4万年前に戻るまでネアンデルタール人に取って代わられた[447]。この同定は2020年の研究によって否定された [448]。考古学的証拠は、ネアンデルタール人が約10万年前に近東で約6~5万年前まで現生人類を追い出していたことを示唆している[100]。 原因 現生人類 ホモ・エレクトス(黄色)、ネアンデルタール人(黄土色)、現生人類(赤色)の相次ぐ離散。 歴史的には、現生人類の技術はネアンデルタール人の技術よりもはるかに優れており、より効率的な武器や自給自足戦略を持っていると見なされており、ネアン デルタール人は競争に勝てなかったために絶滅しただけであった[19]。 ネアンデルタール人と現生人類の内殖の発見は、多地域仮説の復活を引き起こした。多地域仮説では、現在の全人類の遺伝的構成は、世界中に分散している人類 のいくつかの異なる集団間の複雑な遺伝的接触の結果である。このモデルによれば、ネアンデルタール人やその他の最近の古人類は、単に現代人のゲノムに同化 されただけであり、つまり事実上繁殖されて絶滅したのである[19]。現代人は、ヨーロッパにおいて約3,000年から5,000年の間ネアンデルタール 人と共存していた[449]。 気候変動 ネアンデルタール人の最終的な絶滅は、季節性の激しい時期であったハインリッヒ事象4と一致している。その後のハインリッヒ事象は、ヨーロッパ人の集団が 崩壊した際の大規模な文化の転換とも関連している。 [20][21]この気候変動は、以前の寒冷スパイクのようにネアンデルタール人の人口を減少させたかもしれないが、その代わりにこれらの地域は移住して きた人類によって再び人口が増加し、ネアンデルタール人の絶滅につながった[450]。イベリア半島南部では、H4の間にネアンデルタール人の人口が減少 し、それに関連してアルテミシアが支配する砂漠草原が増殖したという証拠がある[451]。 イタリア半島上の赤い斑点が中央ロシアとエーゲ海とイオニア海を越えて東に伸びていることを示すヨーロッパの地図。 約4万年前のカンパニア火山噴火の際の堆積物の分散 ネアンデルタール人とその祖先は、何十万年もの間ヨーロッパに居住し、いくつかの氷河期を生き延びてきた。 [279]また、ネアンデルタール人の個体数が他の要因によって既に減少していたと思われる4万年前頃に、イタリアのカンパニア・イグニンブライト噴火が 1年間2~4℃(3.6~7.2°F)の冷却をもたらし、さらに数年間酸性雨をもたらしたため、ネアンデルタール人の最終的な絶滅につながった可能性があ ると提唱されている[22][453]。 病気 現生人類がアフリカの病気をネアンデルタール人に持ち込み、絶滅の一因となった可能性がある。また、遺伝的多様性が低いために、これらの新しい病気に対す る免疫力を自然に持つネアンデルタール人が少なかった可能性もある(「病原体耐性の差」仮説)。しかし、ネアンデルタール人は現代人と比べて、適応免疫系 に関連する12の主要組織適合複合体(MHC)遺伝子の遺伝的多様性が同等かそれ以上であったことから、このモデルには疑問が投げかけられている [24]。 低人口と近親交配の不況が不適応な先天性欠損を引き起こし、それが彼らの衰退(突然変異によるメルトダウン)の一因となった可能性がある[243]。 20世紀末のニューギニアでは、人肉食による葬儀の習慣のために、フォア族は伝達性海綿状脳症、特に脳組織から発見されたプリオンの摂取によって伝播する 強毒性疾患であるクルによって壊滅的な打撃を受けた。しかし、PRNP遺伝子の129変異体を持つ個体には、もともとプリオンに対する免疫があった。この 遺伝子を研究した結果、129型が現生人類の間に広く存在していることが発見された。ネアンデルタール人はある程度カニバリズムを実践し、現生人類と共存 していたことが知られているため、イギリスの古人類学者サイモン・アンダーダウンは、現生人類がクルのような海綿状疾患をネアンデルタール人に感染させ、 ネアンデルタール人には129変異体が存在しなかったと思われるため、すぐに死滅させたと推測した[23][454]。 |
| https://en.wikipedia.org/wiki/Neanderthal |
|
| The Neanderthal genome project is an effort of a group of scientists to sequence the Neanderthal genome, founded in July 2006. It was initiated by 454 Life Sciences, a biotechnology company based in Branford, Connecticut in the United States and is coordinated by the Max Planck Institute for Evolutionary Anthropology in Germany. In May 2010 the project published their initial draft of the Neanderthal genome (Vi33.16, Vi33.25, Vi33.26) based on the analysis of four billion base pairs of Neanderthal DNA. The study determined that some mixture of genes occurred between Neanderthals and anatomically modern humans and presented evidence that elements of their genome remain in modern humans outside Africa.[1][2][3] In December 2013, a high coverage genome of a Neanderthal was reported for the first time. DNA was extracted from a toe fragment from a female Neanderthal researchers have dubbed the "Altai Neandertal". It was found in Denisova Cave in the Altai Mountains of Siberia and is estimated to be 50,000 years old.[4][5] Findings The researchers recovered ancient DNA of Neanderthals by extracting the DNA from the femur bones of three 38,000 year-old female Neanderthal specimens from Vindija Cave, Croatia, and other bones found in Spain, Russia, and Germany.[6] Only about half a gram of the bone samples (or 21 samples each 50–100 mg[1]) was required for the sequencing, but the project faced many difficulties, including the contamination of the samples by the bacteria that had colonized the Neanderthal's body and humans who handled the bones at the excavation site and at the laboratory.[7]  Svante Pääbo, director of the Department of Genetics at the Max Planck Institute for Evolutionary Anthropology and head of its Neanderthal genome project In February 2009, the Max Planck Institute's team led by Svante Pääbo announced that they had completed the first draft of the Neanderthal genome.[7] An early analysis of the data suggested in "the genome of Neanderthals, a human species driven to extinction" "no significant trace of Neanderthal genes in modern humans".[8] New results suggested that some adult Neanderthals were lactose intolerant.[9] On the question of potentially cloning a Neanderthal, Pääbo commented, "Starting from the DNA extracted from a fossil, it is and will remain impossible."[7] In May 2010, the project released a draft of their report on the sequenced Neanderthal genome. Contradicting the results discovered while examining mitochondrial DNA (mtDNA), they demonstrated a range of genetic contribution to non-African modern humans ranging from 1% to 4%. From their Homo sapiens samples in Eurasia (French, Han Chinese and Papuan) the authors stated that it is likely that interbreeding occurred in the Levant before Homo sapiens migrated into Europe.[10] This finding is disputed because of the paucity of archeological evidence supporting their statement. The fossil evidence does not conclusively place Neanderthals and modern humans in close proximity at this time and place.[11] According to preliminary sequences from 2010, 99.7% of the nucleotide sequences of the modern human and Neanderthal genomes are identical, compared to humans sharing around 98.8% of sequences with the chimpanzee.[12] (For some time, studies concerning the commonality between chimps and humans modified the commonality of 99% to a commonality of only 94%, showing that the genetic gap between humans and chimpanzees was far larger than originally thought,[13][14] but more recent knowledge states the difference between humans, chimpanzees, and bonobos at just about 1.0–1.2% again.[15][16]) Additionally, in 2010, the discovery and analysis of mtDNA from the Denisova hominin in Siberia revealed that it differed from that of modern humans by 385 bases (nucleotides) in the mtDNA strand out of approximately 16,500, whereas the difference between modern humans and Neanderthals is around 202 bases. In contrast, the difference between chimpanzees and modern humans is approximately 1,462 mtDNA base pairs. Analysis of the specimen's nuclear DNA was then still under way and expected to clarify whether the find is a distinct species.[17][18] Even though the Denisova hominin's mtDNA lineage predates the divergence of modern humans and Neanderthals, coalescent theory does not preclude a more recent divergence date for her nuclear DNA. A rib fragment from the partial skeleton of a Neanderthal infant found in the Mezmaiskaya cave in the northwestern foothills of the Caucasus Mountains was radiocarbon-dated in 1999 to 29,195±965 B.P., and therefore belonging to the latest lived Neanderthals. Ancient DNA recovered for a mtDNA sequence showed 3.48% divergence from that of the Feldhofer Neanderthal, some 2,500 km to the west in Germany and in 2011 Phylogenetic analysis placed the two in a clade distinct from modern humans, suggesting that their mtDNA types have not contributed to the modern human mtDNA pool.[19] In 2015, Israel Hershkovitz of Tel Aviv University reported that a skull found in a cave in northern Israel, is "probably a woman, who lived and died in the region about 55,000 years ago, placing modern humans there and then for the first time ever", pointing to a potential time and location when modern humans first interbred with Neanderthals.[20] In 2016, the project found that Neanderthals bred with modern humans multiple times, and that Neanderthals interbred with Denisovans only once, as evidenced in the genome of modern-day Melanesians.[21] In 2006, two research teams working on the same Neanderthal sample published their results, Richard Green and his team in Nature,[22] and James Noonan's team in Science.[23] The results were received with some scepticism, mainly surrounding the issue of a possible admixture of Neanderthals into the modern human genome.[24] In 2006, Richard Green's team had used a then new sequencing technique developed by 454 Life Sciences that amplifies single molecules for characterization and obtained over a quarter of a million unique short sequences ("reads"). The technique delivers randomly located reads, so that sequences of interest – genes that differ between modern humans and Neanderthals – show up at random as well. However, this form of direct sequencing destroys the original sample so to obtain new reads more samples must be destructively sequenced.[25] Noonan's team, led by Edward Rubin, used a different technique, one in which the Neanderthal DNA is inserted into bacteria, which make multiple copies of a single fragment. They demonstrated that Neanderthal genomic sequences can be recovered using a metagenomic library-based approach. All of the DNA in the sample is "immortalized" into metagenomic libraries. A DNA fragment is selected, then propagated in microbes. The resulting Neanderthal DNA sequences can then be sequenced or specific sequences can be studied.[25] Overall, their results were remarkably similar. One group suggested there was a hint of mixing between human and Neanderthal genomes, while the other found none, but both teams recognized that the data set was not large enough to give a definitive answer.[24] The publication by Noonan, and his team revealed Neanderthal DNA sequences matching chimpanzee DNA, but not modern human DNA, at multiple locations, thus enabling the first accurate calculation of the date of the most recent common ancestor of H. sapiens and H. neanderthalensis. The research team estimates the most recent common ancestor of their H. neanderthalensis samples and their H. sapiens reference sequence lived 706,000 years ago (divergence time), estimating the separation of the human and Neanderthal ancestral populations to 370,000 years ago (split time). "Our analyses suggest that on average the Neanderthal genomic sequence we obtained and the reference human genome sequence share a most recent common ancestor ~706,000 years ago, and that the human and Neanderthal ancestral populations split ~370,000 years ago, before the emergence of anatomically modern humans." — Noonan et al. (2006)[23] Based on the analysis of mitochondrial DNA, the split of the Neanderthal and H. sapiens lineages is estimated to date to between 760,000 and 550,000 years ago (95% CI).[26] Mutations of the speech-related gene FOXP2 identical to those in modern humans were discovered in Neanderthal DNA from the El Sidrón 1253 and 1351c specimens,[27] suggesting Neanderthals might have shared some basic language capabilities with modern humans.[9] |
ネアンデルタール人ゲノムプロジェクトは、ネアンデルタール人のゲノムを解読しようとする科学者グループによる取り組みで、2006年7月に発足した。 米国コネチカット州ブランフォードに本社を置くバイオテクノロジー企業454ライフサイエンス社が主導し、ドイツのマックス・プランク進化人類学研究所が コーディネートしている。2010年5月、このプロジェクトは40億塩基対のネアンデルタール人DNAの解析に基づくネアンデルタール人ゲノムの初期草稿 (Vi33.16、Vi33.25、Vi33.26)を発表した。この研究では、ネアンデルタール人と解剖学的に現生人類の間で遺伝子の混合が起こったと 断定し、アフリカ以外の現生人類にもネアンデルタール人のゲノムの要素が残っているという証拠を提示した[1][2][3]。 2013年12月、ネアンデルタール人の高カバレッジゲノムが初めて報告された。DNAは、研究者たちが「アルタイ・ネアンデルタール」と呼んでいる女性 のネアンデルタール人の足の指の断片から抽出された。これはシベリアのアルタイ山脈にあるデニソワ洞窟で発見されたもので、50,000年前のものと推定 されている[4][5]。 発見 研究者たちは、クロアチアのヴィンディヤ洞窟から発見された3万8000年前の女性ネアンデルタール人標本3体の大腿骨と、スペイン、ロシア、ドイツで発 見されたその他の骨からDNAを抽出し、ネアンデルタール人の古代DNAを回収した。 [しかし、ネアンデルタール人の体に住み着いたバクテリアによるサンプルの汚染や、発掘現場や研究所で骨を扱った人間による汚染など、このプロジェクトは 多くの困難に直面した[7]。 マックス・プランク進化人類学研究所の遺伝学部長であり、ネアンデルタール人ゲノムプロジェクトの責任者であるスヴァンテ・ペーボ(Svante Pääbo)氏。 2009年2月、スヴァンテ・ペーボ率いるマックス・プランク研究所のチームは、ネアンデルタール人ゲノムの最初の草稿を完成させたと発表した[7]。初 期のデータ解析では、「絶滅に追いやられた人類種であるネアンデルタール人のゲノム」には「現代人にネアンデルタール人の遺伝子の有意な痕跡はない」こと が示唆されていた[8]。 [8]。ネアンデルタール人のクローンを作る可能性について、ペーボは「化石から抽出したDNAから始めることは不可能であり、今後も不可能である」とコ メントした[7]。 2010年5月、プロジェクトは、解読されたネアンデルタール人ゲノムに関する報告書の草稿を発表した。ミトコンドリアDNA(mtDNA)の調査で発見 された結果とは対照的に、アフリカ以外の現生人類への遺伝的寄与は1%から4%の範囲に及ぶことが示された。著者はユーラシア大陸のホモ・サピエンス(フ ランス人、漢民族、パプア人)のサンプルから、ホモ・サピエンスがヨーロッパに移動する前にレバントで交雑が起こった可能性が高いと述べている[10]。 2010年に発表された予備的な塩基配列によると、現生人類とネアンデルタール人のゲノムの塩基配列の99.7%が同一であり、チンパンジーの塩基配列の 98.8%が同一である。 [12](しばらくの間、チンパンジーとヒトの共通性に関する研究は、99%の共通性をわずか94%の共通性に修正し、ヒトとチンパンジーの間の遺伝的 ギャップが当初考えられていたよりもはるかに大きいことを示していた[13][14]が、より最近の知見では、ヒト、チンパンジー、ボノボの間の差はわず か1.0-1.2%程度であるとされている[15][16])。 さらに2010年、シベリアのデニソワホミニンからmtDNAが発見され、解析された結果、mtDNA鎖の約16,500塩基のうち385塩基(ヌクレオ チド)が現生人類と異なることが明らかになったが、現生人類とネアンデルタール人の違いは約202塩基である。一方、チンパンジーと現生人類のmtDNA 塩基対の差は約1,462である。デニソワホミニンのmtDNAの系統が現生人類とネアンデルタール人の分岐よりも先であったとしても、核DNAの分岐年 代がより新しいことを合体説は否定していない[17][18]。 コーカサス山脈の北西麓にあるメズマイスカヤ洞窟で発見されたネアンデルタール人の幼児の部分骨格から採取された肋骨断片は、1999年に放射性炭素年代 測定が行われ、紀元前29,195±965年であった。mtDNA配列のために回収された古代のDNAは、西へ約2,500km離れたドイツのフェルド ホーファー・ネアンデルタール人のものと3.48%の分岐を示し、2011年の系統解析では、この2人は現生人類とは異なるクレードに位置づけられ、彼ら のmtDNA型は現生人類のmtDNAプールに寄与していないことが示唆された[19]。 2015年、テルアビブ大学のイスラエル・ハーシュコヴィッツは、イスラエル北部の洞窟で発見された頭蓋骨が、「おそらく女性で、約55,000年前にこ の地域に住んで死亡し、現生人類をそこに置き、そして史上初めて」、現生人類がネアンデルタール人と初めて交雑した時期と場所の可能性を指摘していると報 告した[20]。 2016年、このプロジェクトは、ネアンデルタール人が現生人類と複数回交配し、ネアンデルタール人がデニソワ人と1回だけ交配したことを、現代のメラネシア人のゲノムから明らかにした[21]。 2006年、同じネアンデルタール人のサンプルを研究していた2つの研究チームが、リチャード・グリーンと彼のチームは『ネイチャー』誌に、ジェームズ・ヌーナンのチームは『サイエンス』誌にその結果を発表した[22]。 2006年、リチャード・グリーンの研究チームは、454ライフサイエンス社が開発した、単一分子を増幅して特性解析を行う当時としては新しいシークエン シング技術を使用し、25万以上のユニークな短い配列(「リード」)を得た。この技術では、ランダムに配置されたリードが得られるため、興味のある配列 (現代人とネアンデルタール人の間で異なる遺伝子)もランダムに現れる。しかし、このダイレクトシークエンシングは元のサンプルを破壊してしまうため、新 しいリードを得るためには、より多くのサンプルを破壊的にシークエンシングしなければならない[25]。 エドワード・ルービン率いるヌーナンの研究チームは、ネアンデルタール人のDNAをバクテリアに挿入し、ひとつの断片から複数のコピーを作るという別の手 法を用いた。彼らは、メタゲノムライブラリーを用いたアプローチで、ネアンデルタール人のゲノム配列を回収できることを実証した。サンプル中のDNAはす べてメタゲノムライブラリーに「不死化」される。DNA断片を選択し、微生物の中で増殖させる。こうして得られたネアンデルタール人のDNA配列は、塩基 配列を決定したり、特定の配列を研究したりすることができる[25]。 全体的に、その結果は驚くほど似ていた。あるグループはヒトとネアンデルタール人のゲノムの間に混血のヒントがあることを示唆し、もう一方は何も発見しなかったが、両チームともデータセットが決定的な答えを出すのに十分な大きさではないことを認識していた[24]。 ヌーナンらの研究チームは、ネアンデルタール人のDNA配列がチンパンジーのDNAと一致し、現代人のDNAとは一致しないことを複数の場所で明らかにし た。研究チームは、H.ネアンデルタール人サンプルとH.サピエンス参照配列の最近の共通祖先は70万6千年前(分岐時間)に生きていたと推定しており、 ヒトとネアンデルタール人の祖先集団の分離は37万年前(分裂時間)と推定している。 「我々の分析によれば、我々が得たネアンデルタール人のゲノム配列と参照ヒトゲノム配列は、平均して〜706,000年前に直近の共通祖先を共有してお り、ヒトとネアンデルタール人の祖先集団は、解剖学的に現生人類が出現する前の〜370,000年前に分裂したことが示唆される」 - ヌーナンら(2006)[23]。 ミトコンドリアDNAの分析に基づくと、ネアンデルタール人とH.サピエンスの系統の分裂は76万年から55万年前(95%信頼区間)と推定されている[26]。 エル・シドロン1253号と1351c号の標本から採取されたネアンデルタール人のDNAから、現生人類と同一の音声関連遺伝子FOXP2の変異が発見されており[27]、ネアンデルタール人が現生人類と基本的な言語能力を共有していた可能性を示唆している[9]。 |
| Cro-Magnon – Earliest anatomically modern humans in Europe Neanderthal extinction – Prehistoric event Interbreeding between archaic and modern humans – Evidence of human hybridization during the Paleolithic General: Admixture mapping – Result of interbreeding between two or more previously isolated populations within a species Gene flow – Transfer of genetic variation from one population to another Hybrid – Offspring of cross-species reproduction |
クロマニヨン人 - ヨーロッパ最古の解剖学的現生人類 ネアンデルタール人の絶滅 - 先史時代の出来事 古人類と現生人類の交雑 - 旧石器時代におけるヒトの交雑の証拠 一般的に 混血マッピング - ある種の中で、以前は隔離されていた2つ以上の個体群間の交雑の結果である。 遺伝子の流れ - ある集団から別の集団への遺伝的変異の移動 雑種 - 異種間繁殖の子孫 |
| https://en.wikipedia.org/wiki/Neanderthal_genome_project |
|
| Demographics Further information: Neanderthals in Southwest Asia, Neanderthals in Gibraltar, and List of Neanderthal sites Range  A skull missing most of the left side of the face from the mid-orbit to the teeth Neanderthal skull from Tabun Cave, Israel, at the Israel Museum Pre- and early Neanderthals, living before the Eemian interglacial (130,000 years ago), are poorly known and come mostly from Western European sites. From 130,000 years ago onwards, the quality of the fossil record increases dramatically with classic Neanderthals, who are recorded from Western, Central, Eastern and Mediterranean Europe,[15] as well as Southwest, Central and Northern Asia up to the Altai Mountains in southern Siberia. Pre- and early Neanderthals, on the other hand, seem to have continuously occupied only France, Spain and Italy, although some appear to have moved out of this "core-area" to form temporary settlements eastward (although without leaving Europe). Nonetheless, southwestern France has the highest density of sites for pre-, early and classic Neanderthals.[160] The Neanderthals were the first human species to permanently occupy Europe as the continent was only sporadically occupied by earlier humans.[161]Neanderthals are technically or in some aspect, the true and only indigenous humans populace of Europe. The southernmost find was recorded at Shuqba Cave, Levant;[162] reports of Neanderthals from the North African Jebel Irhoud[163] and Haua Fteah[164] have been reidentified as H. sapiens. Their easternmost presence is recorded at Denisova Cave, Siberia 85°E; the southeast Chinese Maba Man, a skull, shares several physical attributes with Neanderthals, although these may be the result of convergent evolution rather than Neanderthals extending their range to the Pacific Ocean.[165] The northernmost bound is generally accepted to have been 55°N, with unambiguous sites known between 50–53°N, although this is difficult to assess because glacial advances destroy most human remains, and palaeoanthropologist Trine Kellberg Nielsen has argued that a lack of evidence of Southern Scandinavian occupation is (at least during the Eemian interglacial) due to the former explanation and a lack of research in the area.[166][167] Middle Palaeolithic artefacts have been found up to 60°N on the Russian plains,[168][169][170] but these are more likely attributed to modern humans.[171] A 2017 study claimed the presence of Homo at the 130,000-year-old Californian Cerutti Mastodon site in North America,[172] but this is largely considered implausible.[173][174][175] It is unknown how the rapidly fluctuating climate of the last glacial period (Dansgaard–Oeschger events) impacted Neanderthals, as warming periods would produce more favourable temperatures but encourage forest growth and deter megafauna, whereas frigid periods would produce the opposite.[176] However, Neanderthals may have preferred a forested landscape.[71] Stable environments with mild mean annual temperatures may have been the most suitable Neanderthal habitats.[177] Populations may have peaked in cold but not extreme intervals, such as marine isotope stages 8 and 6 (respectively, 300,000 and 191,000 years ago during the Saale glaciation). It is possible their range expanded and contracted as the ice retreated and grew, respectively, to avoid permafrost areas, residing in certain refuge zones during glacial maxima.[176] In 2021, Israeli anthropologist Israel Hershkovitz and colleagues suggested the 140- to 120,000-year-old Israeli Nesher Ramla remains, which feature a mix of Neanderthal and more ancient H. erectus traits, represent one such source population which recolonised Europe following a glacial period.[178] 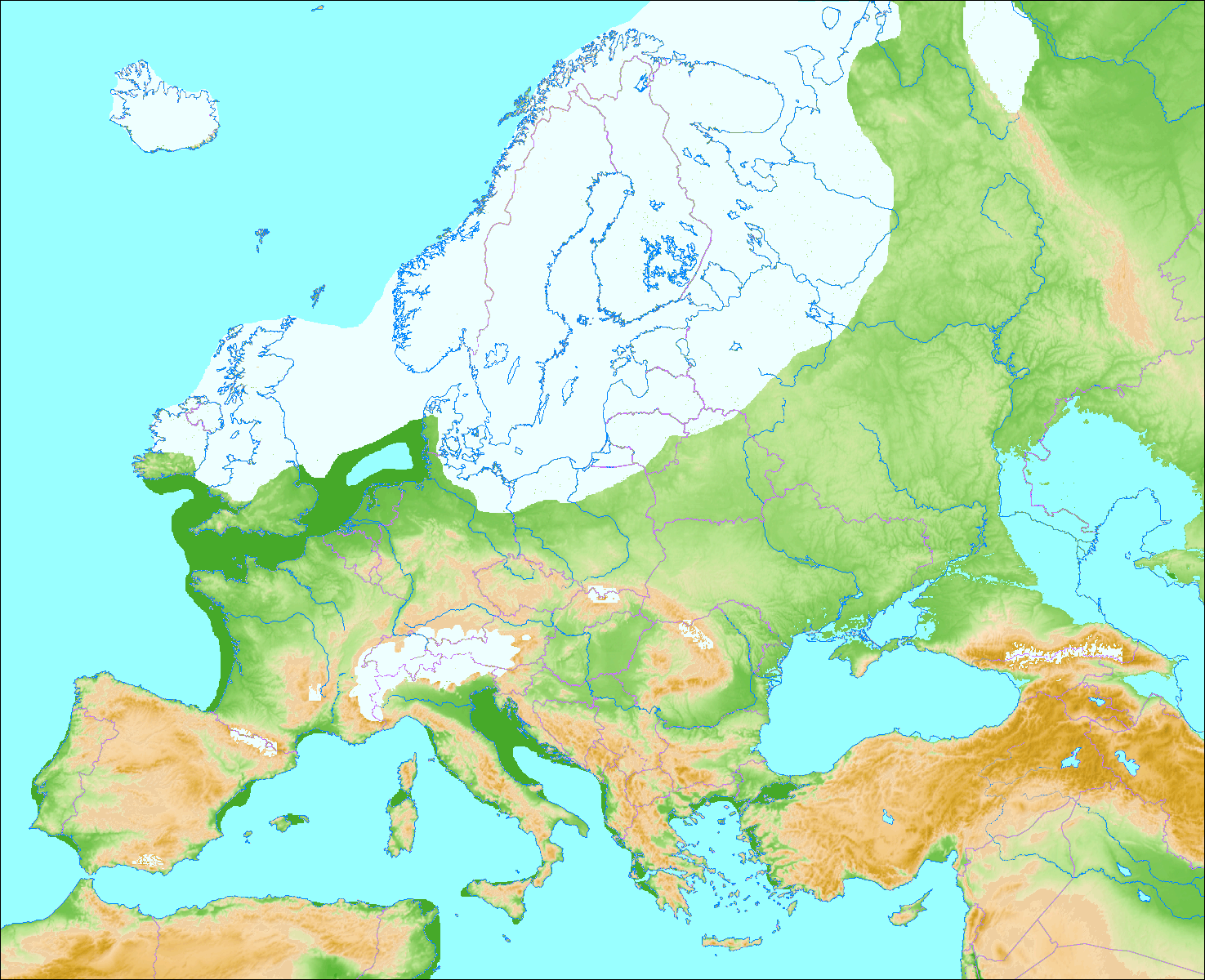 Map of Europe during the Würm glaciation 70–20,000 years ago Population Like modern humans, Neanderthals probably descended from a very small population with an effective population—the number of individuals who can bear or father children—of 3,000 to 12,000 approximately. However, Neanderthals maintained this very low population, proliferating weakly harmful genes due to the reduced effectivity of natural selection.[26][179] Various studies, using mtDNA analysis, yield varying effective populations,[176] such as about 1,000 to 5,000;[179] 5,000 to 9,000 remaining constant;[180] or 3,000 to 25,000 steadily increasing until 52,000 years ago before declining until extinction.[28] Archaeological evidence suggests that there was a tenfold increase in the modern human population in Western Europe during the period of the Neanderthal/modern human transition,[181] and Neanderthals may have been at a demographic disadvantage due to a lower fertility rate, a higher infant mortality rate, or a combination of the two.[182] Estimates giving a total population in the higher tens of thousands[138] are contested.[179] A consistently low population may be explained in the context of the "Boserupian Trap": a population's carrying capacity is limited by the amount of food it can obtain, which in turn is limited by its technology. Innovation increases with population, but if the population is too low, innovation will not occur very rapidly and the population will remain low. This is consistent with the apparent 150,000 year stagnation in Neanderthal lithic technology.[176] In a sample of 206 Neanderthals, based on the abundance of young and mature adults in comparison to other age demographics, about 80% of them above the age of 20 died before reaching 40. This high mortality rate was probably due to their high-stress environment.[25] However, it has also been estimated that the age pyramids for Neanderthals and contemporary modern humans were the same.[176] Infant mortality was estimated to have been very high for Neanderthals, about 43% in northern Eurasia.[183] 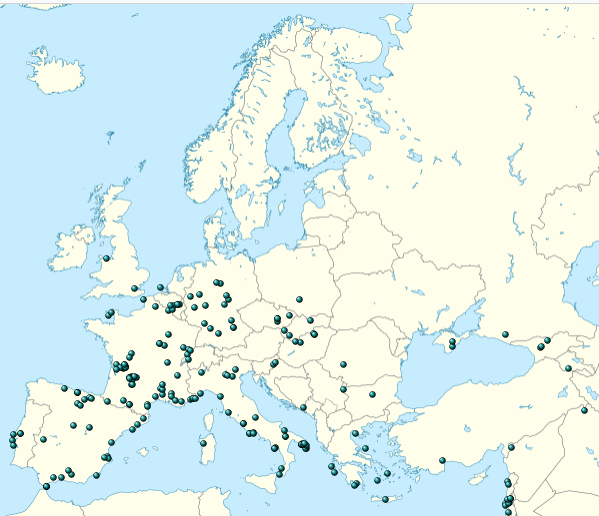 Locations of Neanderthal finds in Europe and the Levant. https://en.wikipedia.org/wiki/Neanderthal |
人口 さらなる情報 南西アジアのネアンデルタール人、ジブラルタルのネアンデルタール人、ネアンデルタール人遺跡のリスト 範囲 顔の左側、正中軌道から歯にかけての大部分を失った頭蓋骨。 イスラエル、タブン洞窟出土のネアンデルタール人頭蓋骨(イスラエル博物館所蔵)。 エマ紀間氷期(13万年前)以前のネアンデルタール人はあまり知られておらず、ほとんどが西ヨーロッパの遺跡から得られている。130,000年前以降、 古典的なネアンデルタール人の化石記録の質は飛躍的に高まり、西ヨーロッパ、中央ヨーロッパ、東ヨーロッパ、地中海ヨーロッパ[15]、さらに南西アジ ア、中央アジア、北アジアからシベリア南部のアルタイ山脈まで記録されている。一方、ネアンデルタール人以前および初期は、フランス、スペイン、イタリア にのみ継続して居住していたようであるが、一部はこの「中心地域」から移動して、(ヨーロッパを離れることなく)東方に一時的な集落を形成したようであ る。それにもかかわらず、フランス南西部には、ネアンデルタール人以前、初期、および古典的なネアンデルタール人の遺跡が最も密集している[160]。ネ アンデルタール人は、ヨーロッパ大陸を永続的に占領した最初の人類種であった。 北アフリカのジェベル・イルフード[163]とハウア・フテア[164]で発見されたネアンデルタール人の報告は、サピエンスと再同定されている。ネアン デルタール人の最東端は、東経85度のシベリアのデニソワ洞窟に記録されている。中国南東部のマバ人(頭骨)は、ネアンデルタール人といくつかの身体的特 徴を共有しているが、これらはネアンデルタール人が太平洋まで範囲を広げたというよりも、収斂進化の結果である可能性がある。 [165] 最北端は北緯55度であると一般的に受け入れられており、北緯50度から53度の間に明確な遺跡が確認されているが、氷河期の進行によってほとんどの人骨 が破壊されているため、これを評価するのは困難である。古人類学者のトリーネ・ケルベルグ・ニールセンは、南スカンジナビア人が居住していた証拠がないの は(少なくともエマ紀間氷期の間)、前者の説明とこの地域の調査不足によるものだと主張している。 [166][167]中旧石器時代の遺物はロシアの平原で北緯60度まで発見されている[168][169][170]が、これらは現生人類に起因する可 能性が高い[171]。 2017年の研究では、北米の13万年前のカリフォルニアのセルッティ・マストドン遺跡にホモが存在すると主張している[172]が、これはほとんどあり 得ないと考えられている[173][174][175]。 最終氷期の気候の急激な変動(ダンスガード・オエシュガー・イベント)がネアンデルタール人にどのような影響を与えたかは不明である。温暖な時期はより好 ましい気温をもたらすが、森林の成長を促し、巨大動物を抑止する。 [海洋同位体第8段階と第6段階(それぞれ30万年前と19万1,000年前のザーレ氷河期)のように、極端ではないが寒冷な時期に個体数がピークに達し た可能性がある。氷河期の最盛期には、永久凍土地域を避け、特定の避難地帯に居住していた。2021年、イスラエルの人類学者イスラエル・ハーシュコ ヴィッツと同僚は、ネアンデルタール人とより古代のH.エレクトゥスの形質が混在している14~12万年前のイスラエルのネッシャー・ラムラの遺跡が、氷 河期に続いてヨーロッパを再植民地化したそのような源集団の一つであることを示唆した[178]。 7~2万年前のヴュルム氷河期のヨーロッパ地図 人口 現生人類と同様に、ネアンデルタール人はおそらく非常に小さな集団の子孫であり、有効人口(子供を産んだり父親になったりできる個体数)はおよそ 3,000人から12,000人であった。しかし、ネアンデルタール人はこの非常に低い集団を維持し、自然淘汰の効果が減少したために、弱く有害な遺伝子 を増殖させていた[26][179]。mtDNA分析を用いた様々な研究によって、有効人口は様々であり[176]、例えば、約1,000から 5,000、[179]5,000から9,000は一定、[180]あるいは3,000から25,000は52,000年前まで着実に増加し、その後絶滅 するまで減少した。 [ネアンデルタール人と現生人類の移行期に、西ヨーロッパでは現生人類の人口が10倍に増加したことを考古学的証拠が示唆している[181]。 [182]総人口を数万人以上とする推定[138]については異論がある[179]。一貫して人口が少ないことは、「ボセループの罠」という文脈で説明す ることができるかもしれない。イノベーションは人口とともに増加するが、人口が少なすぎると、イノベーションはあまり急速に起こらず、人口は低いままであ る。このことは、ネアンデルタール人の石器技術が明らかに15万年間停滞していることと一致している[176]。 206人のネアンデルタール人のサンプルにおいて、他の年齢層と比較した若年層と壮年層の多さに基づくと、20歳以上のネアンデルタール人の約80%が 40歳に達する前に死亡している。この高い死亡率は、おそらく彼らの高ストレス環境によるものであった[25]。しかし、ネアンデルタール人と現代の現代 人の年齢ピラミッドは同じであったと推定されている[176]。ネアンデルタール人の乳児死亡率は非常に高く、ユーラシア北部では約43%であったと推定 されている[183]。 |
| Anatomy Main article: Neanderthal anatomy Build 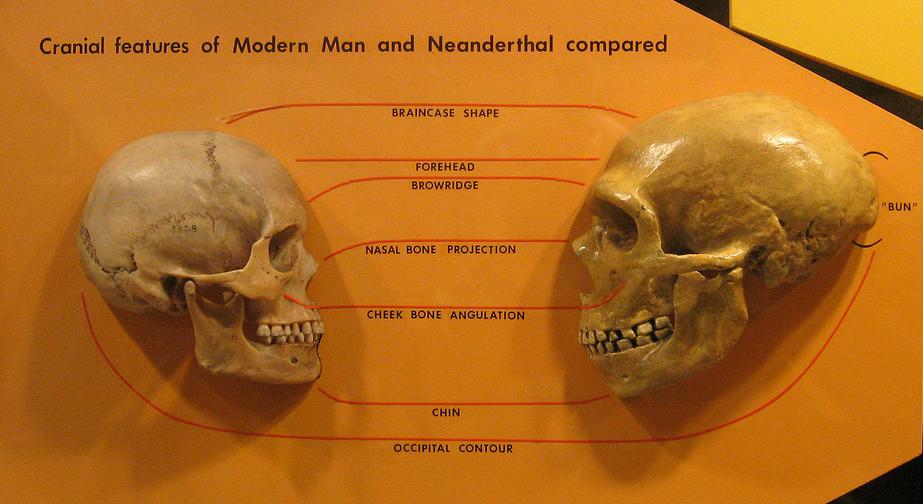 A human skull on the left facing a reconstructed Neanderthal skull on the right, emphasising the difference in braincase shape (more cranial length in Neanderthal), shorter forehead ratio, more defined brow ridge, larger nasal bone projection, pinned-back cheekbone angulation, straighter angled chin, and an occipital bun Comparisons of a modern Eurasian male example (left) and a Neanderthal (right) skull reconstruction at the Cleveland Museum of Natural History 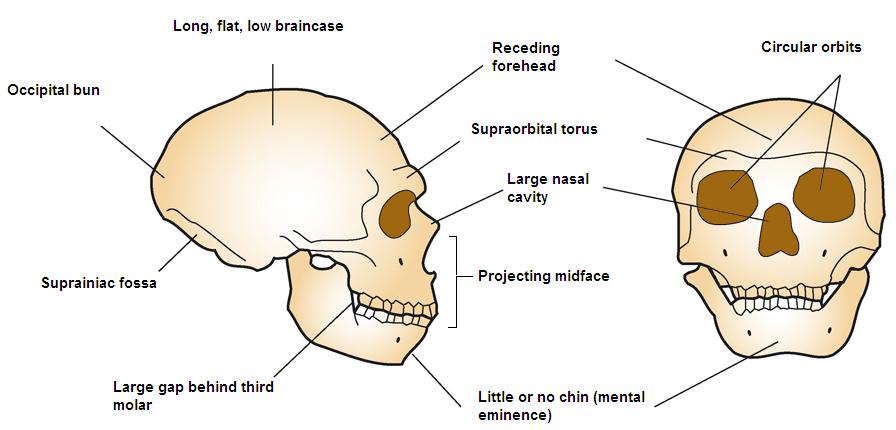 Front and side view diagram of Neanderthal skull reconstruction emphasising large circular orbits, straightened chin, projecting nasal bridge, large brow ridge, receded forehead, long topped braincase, occipital bun, fossa, and a large gap behind the third molar Neanderthal skull features Neanderthals had more robust and stockier builds than typical modern humans,[75] wider and barrel-shaped rib cages; wider pelvises;[30][184] and proportionally shorter forearms and forelegs.[71][185] Based on 45 Neanderthal long bones from 14 men and 7 women, the average height was 164 to 168 cm (5 ft 5 in to 5 ft 6 in) for males and 152 to 156 cm (5 ft 0 in to 5 ft 1 in) for females.[75] For comparison, the average height of 20 males and 10 females Upper Palaeolithic humans is, respectively, 176.2 cm (5 ft 9.4 in) and 162.9 cm (5 ft 4.1 in), although this decreases by 10 cm (4 in) nearer the end of the period based on 21 males and 15 females;[186] and the average in the year 1900 was 163 cm (5 ft 4 in) and 152.7 cm (5 ft 0 in), respectively.[187] The fossil record shows that adult Neanderthals varied from about 147.5 to 177 cm (4 ft 10 in to 5 ft 10 in) in height, although some may have grown much taller (73.8 to 184.8 cm based on footprint length and from 65.8 to 189.3 cm based on footprint width).[188] For Neanderthal weight, samples of 26 specimens found an average of 77.6 kg (171 lb) for males and 66.4 kg (146 lb) for females.[189] Using 76 kg (168 lb), the body mass index for Neanderthal males was calculated to be 26.9–28.2, which in modern humans correlates to being overweight. This indicates a very robust build.[75] The Neanderthal LEPR gene concerned with storing fat and body heat production is similar to that of the woolly mammoth, and so was likely an adaptation for cold climate.[72]  Neanderthal hunters depicted in the Gallo-Roman Museum, Tongeren The neck vertebrae of Neanderthals are thicker from the front to the rear and transversely than those of (most) modern humans, leading to stability, possibly to accommodate a different head shape and size.[190] Although the Neanderthal thorax (where the ribcage is) was similar in size to modern humans, the longer and straighter ribs would have equated to a widened mid-lower thorax and stronger breathing in the lower thorax, which are indicative of a larger diaphragm and possibly greater lung capacity.[184][191][192] The lung capacity of Kebara 2 was estimated to have been 9.04 L (2.39 US gal), compared to the average human capacity of 6 L (1.6 US gal) for males and 4.7 L (1.2 US gal) for females. The Neanderthal chest was also more pronounced (expanded front-to-back, or antero-posteriorly). The sacrum (where the pelvis connects to the spine) was more vertically inclined, and was placed lower in relation to the pelvis, causing the spine to be less curved (exhibit less lordosis) and to fold in on itself somewhat (to be invaginated). In modern populations, this condition affects just a proportion of the population, and is known as a lumbarised sacrum.[193] Such modifications to the spine would have enhanced side-to-side (mediolateral) flexion, better supporting the wider lower thorax. It is claimed by some that this feature would be normal for all Homo, even tropically-adapted Homo ergaster or erectus, with the condition of a narrower thorax in most modern humans being a unique characteristic.[184] Body proportions are usually cited as being "hyperarctic" as adaptations to the cold, because they are similar to those of human populations which developed in cold climates[194]—the Neanderthal build is most similar to that of Inuit and Siberian Yupiks among modern humans[195]—and shorter limbs result in higher retention of body heat.[185][194][196] Nonetheless, Neanderthals from more temperate climates—such as Iberia—still retain the "hyperarctic" physique.[197] In 2019, English anthropologist John Stewart and colleagues suggested Neanderthals instead were adapted for sprinting, because of evidence of Neanderthals preferring warmer wooded areas over the colder mammoth steppe, and DNA analysis indicating a higher proportion of fast-twitch muscle fibres in Neanderthals than in modern humans. He explained their body proportions and greater muscle mass as adaptations to sprinting as opposed to the endurance-oriented modern human physique,[71] as persistence hunting may only be effective in hot climates where the hunter can run prey to the point of heat exhaustion (hyperthermia). They had longer heel bones,[198] reducing their ability for endurance running, and their shorter limbs would have reduced moment arm at the limbs, allowing for greater net rotational force at the wrists and ankles, causing faster acceleration.[71] In 1981, American palaeoanthropologist Erik Trinkaus made note of this alternate explanation, but considered it less likely.[185][199] Face Neanderthal man reconstruction, Natural History Museum, London. Neanderthals had less developed chins, sloping foreheads, and longer, broader, more projecting noses. The Neanderthal skull is typically more elongated, but also wider, and less globular than that of most modern humans, and features much more of an occipital bun,[200] or "chignon", a protrusion on the back of the skull, although it is within the range of variation for modern humans who have it. It is caused by the cranial base and temporal bones being placed higher and more towards the front of the skull, and a flatter skullcap.[201] The Neanderthal face is characterised by subnasal[202] as well as mid-facial prognathism, where the zygomatic arches are positioned in a rearward location relative to modern humans, while their maxillary bones and nasal bones are positioned in a more forward direction, by comparison.[203] Neanderthal eyeballs are larger than those of modern humans. One study proposed that this was due to Neanderthals having enhanced visual abilities, at the expense of neocortical and social development.[204] However, this study was rejected by other researchers who concluded that eyeball size does not offer any evidence for the cognitive abilities of Neanderthal or modern humans.[205] The projected Neanderthal nose and paranasal sinuses have generally been explained as having warmed air as it entered the lungs and retained moisture ("nasal radiator" hypothesis);[206] if their noses were wider, it would differ to the generally narrowed shape in cold-adapted creatures, and that it would have been caused instead by genetic drift. Also, the sinuses reconstructed wide are not grossly large, being comparable in size to those of modern humans. However, if sinus size is not an important factor for breathing cold air, then the actual function would be unclear, so they may not be a good indicator of evolutionary pressures to evolve such a nose.[207] Further, a computer reconstruction of the Neanderthal nose and predicted soft tissue patterns shows some similarities to those of modern Arctic peoples, potentially meaning the noses of both populations convergently evolved for breathing cold, dry air.[73] Neanderthals featured a rather large jaw which was once cited as a response to a large bite force evidenced by heavy wearing of Neanderthal front teeth (the "anterior dental loading" hypothesis), but similar wearing trends are seen in contemporary humans. It could also have evolved to fit larger teeth in the jaw, which would better resist wear and abrasion,[206][208] and the increased wear on the front teeth compared to the back teeth probably stems from repetitive use. Neanderthal dental wear patterns are most similar to those of modern Inuit.[206] The incisors are large and shovel-shaped, and, compared to modern humans, there was an unusually high frequency of taurodontism, a condition where the molars are bulkier due to an enlarged pulp (tooth core). Taurodontism was once thought to have been a distinguishing characteristic of Neanderthals which lent some mechanical advantage or stemmed from repetitive use, but was more likely simply a product of genetic drift.[209] The bite force of Neanderthals and modern humans is now thought to be about the same,[206] about 285 N (64 lbf) and 255 N (57 lbf) in modern human males and females, respectively.[210]  Reconstruction of an elderly Neanderthal man Brain The Neanderthal braincase averages 1,640 cm3 (100 cu in) for males and 1,460 cm3 (89 cu in) for females,[77][78] which is significantly larger than the averages for all groups of extant humans;[79] for example, modern European males average 1,362 cm3 (83.1 cu in) and females 1,201 cm3 (73.3 cu in).[211] For 28 modern human specimens from 190,000 to 25,000 years ago, the average was about 1,478 cm3 (90.2 cu in) disregarding sex, and modern human brain size is suggested to have decreased since the Upper Palaeolithic.[212] The largest Neanderthal brain, Amud 1, was calculated to be 1,736 cm3 (105.9 cu in), one of the largest ever recorded in hominids.[78] Both Neanderthal and human infants measure about 400 cm3 (24 cu in).[213] When viewed from the rear, the Neanderthal braincase has lower, wider, rounder appearance than in anatomically modern humans. This characteristic shape is referred to as "en bombe" (bomb-like), and is unique to Neanderthals, with all other hominid species (including most modern humans) generally having narrow and relatively upright cranial vaults, when viewed from behind.[214][215][216][217] The Neanderthal brain would have been characterised by relatively smaller parietal lobes[85] and a larger cerebellum.[85][218] Neanderthal brains also have larger occipital lobes (relating to the classic occurrence of an occipital bun in Neanderthal skull anatomy, as well as the greater width of their skulls), which implies internal differences in the proportionality of brain-internal regions, relative to Homo sapiens, consistent with external measurements obtained with fossil skulls.[204][219] Their brains also have larger temporal lobe poles,[218] wider orbitofrontal cortex,[220] and larger olfactory bulbs,[221] suggesting potential differences in language comprehension and associations with emotions (temporal functions), decision making (the orbitofrontal cortex) and sense of smell (olfactory bulbs). Their brains also show different rates of brain growth and development.[222] Such differences, while slight, would have been visible to natural selection and may underlie and explain differences in the material record in things like social behaviours, technological innovation and artistic output.[18][223] Hair and skin colour The lack of sunlight most likely led to the proliferation of lighter skin in Neanderthals;[224] however, it has been recently claimed that light skin in modern Europeans was not particularly prolific until perhaps the Bronze Age.[225] Genetically, BNC2 was present in Neanderthals, which is associated with light skin colour; however, a second variation of BNC2 was also present, which in modern populations is associated with darker skin colour in the UK Biobank.[224] DNA analysis of three Neanderthal females from southeastern Europe indicates that they had brown eyes, dark skin colour and brown hair, with one having red hair.[226][227] In modern humans, skin and hair colour is regulated by the melanocyte-stimulating hormone—which increases the proportion of eumelanin (black pigment) to phaeomelanin (red pigment)—which is encoded by the MC1R gene. There are five known variants in modern humans of the gene which cause loss-of-function and are associated with light skin and hair colour, and another unknown variant in Neanderthals (the R307G variant) which could be associated with pale skin and red hair. The R307G variant was identified in a Neanderthal from Monti Lessini, Italy, and possibly Cueva del Sidrón, Spain.[228] However, as in modern humans, red was probably not a very common hair colour because the variant is not present in many other sequenced Neanderthals.[224] Metabolism Maximum natural lifespan and the timing of adulthood, menopause and gestation were most likely very similar to modern humans.[176] However, it has been hypothesised, based on the growth rates of teeth and tooth enamel,[229][230] that Neanderthals matured faster than modern humans, although this is not backed up by age biomarkers.[25] The main differences in maturation are the atlas bone in the neck as well as the middle thoracic vertebrae fused about 2 years later in Neanderthals than in modern humans, but this was more likely caused by a difference in anatomy rather than growth rate.[231][232] Generally, models on Neanderthal caloric requirements report significantly higher intakes than those of modern humans because they typically assume Neanderthals had higher basal metabolic rates (BMRs) due to higher muscle mass, faster growth rate and greater body heat production against the cold;[233][234][235] and higher daily physical activity levels (PALs) due to greater daily travelling distances while foraging.[234][235] However, using a high BMR and PAL, American archaeologist Bryan Hockett estimated that a pregnant Neanderthal would have consumed 5,500 calories per day, which would have necessitated a heavy reliance on big game meat; such a diet would have caused numerous deficiencies or nutrient poisonings, so he concluded that these are poorly warranted assumptions to make.[235] Neanderthals may have been more active during dimmer light conditions rather than broad daylight because they lived in regions with reduced daytime hours in the winter, hunted large game (such predators typically hunt at night to enhance ambush tactics), and had large eyes and visual processing neural centres. Genetically, colour blindness (which may enhance mesopic vision) is typically correlated with northern-latitude populations, and the Neanderthals from Vindija Cave, Croatia, had some substitutions in the Opsin genes which could have influenced colour vision. However, the functional implications of these substitutions are inconclusive.[236] Neanderthal-derived alleles near ASB1 and EXOC6 are associated with being an evening person, narcolepsy and day-time napping.[224] Pathology Neanderthals suffered a high rate of traumatic injury, with an estimated 79–94% of specimens showing evidence of healed major trauma, of which 37–52% were severely injured, and 13–19% injured before reaching adulthood.[237] One extreme example is Shanidar 1, who shows signs of an amputation of the right arm likely due to a nonunion after breaking a bone in adolescence, osteomyelitis (a bone infection) on the left clavicle, an abnormal gait, vision problems in the left eye, and possible hearing loss[238] (perhaps swimmer's ear).[239] In 1995, Trinkaus estimated that about 80% succumbed to their injuries and died before reaching 40, and thus theorised that Neanderthals employed a risky hunting strategy ("rodeo rider" hypothesis).[25] However, rates of cranial trauma are not significantly different between Neanderthals and Middle Palaeolithic modern humans (although Neanderthals seem to have had a higher mortality risk),[240] there are few specimens of both Upper Palaeolithic modern humans and Neanderthals who died after the age of 40,[182] and there are overall similar injury patterns between them. In 2012, Trinkaus concluded that Neanderthals instead injured themselves in the same way as contemporary humans, such as by interpersonal violence.[241] A 2016 study looking at 124 Neanderthal specimens argued that high trauma rates were instead caused by animal attacks, and found that about 36% of the sample were victims of bear attacks, 21% big cat attacks, and 17% wolf attacks (totalling 92 positive cases, 74%). There were no cases of hyena attacks, although hyenas still nonetheless probably attacked Neanderthals, at least opportunistically.[242] Such intense predation probably stemmed from common confrontations due to competition over food and cave space, and from Neanderthals hunting these carnivores.[242] 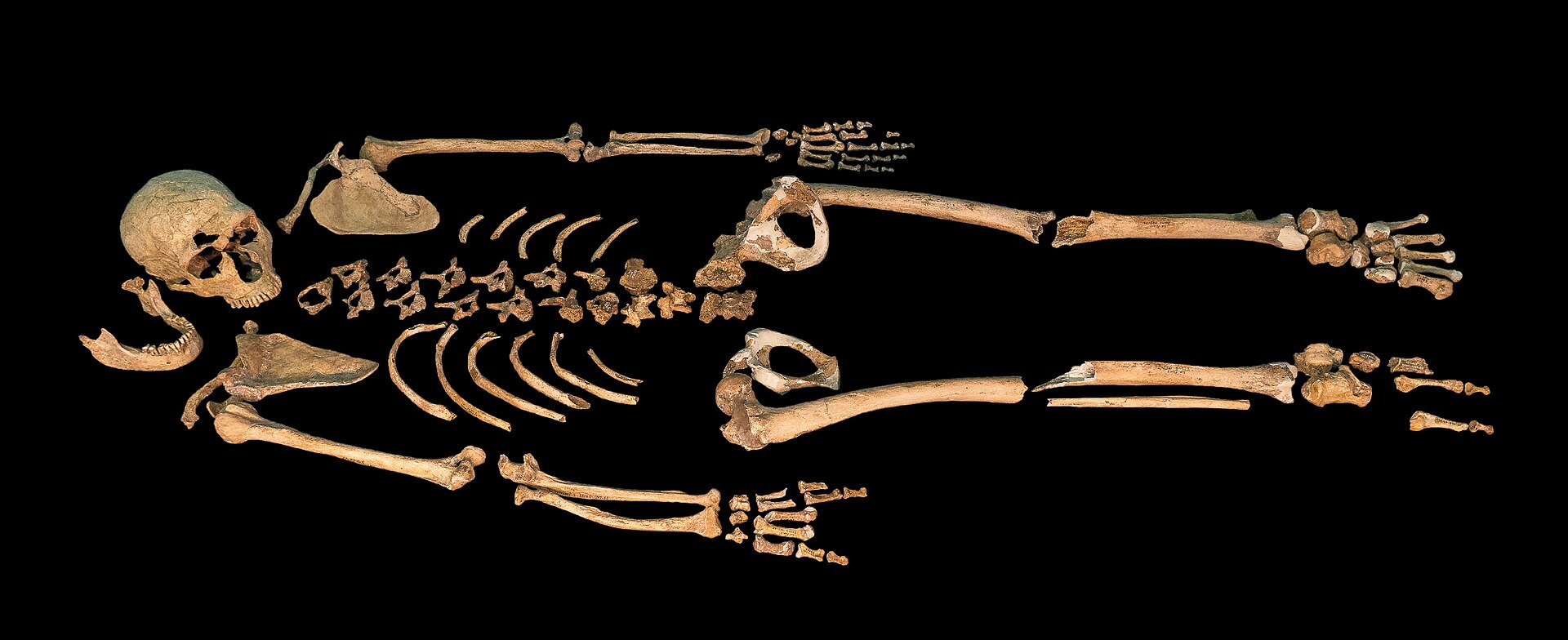 A mostly complete skeleton laid out against a black background horizontally La Ferrassie 1 at the Musée de l'Homme, Paris Low population caused a low genetic diversity and probably inbreeding, which reduced the population's ability to filter out harmful mutations (inbreeding depression). However, it is unknown how this affected a single Neanderthal's genetic burden and, thus, if this caused a higher rate of birth defects than in modern humans.[243] It is known, however, that the 13 inhabitants of Sidrón Cave collectively exhibited 17 different birth defects likely due to inbreeding or recessive disorders.[244] Likely due to advanced age (60s or 70s), La Chapelle-aux-Saints 1 had signs of Baastrup's disease, affecting the spine, and osteoarthritis.[245] Shanidar 1, who likely died at about 30 or 40, was diagnosed with the most ancient case of diffuse idiopathic skeletal hyperostosis (DISH), a degenerative disease which can restrict movement, which, if correct, would indicate a moderately high incident rate for older Neanderthals.[246] Neanderthals were subject to several infectious diseases and parasites. Modern humans likely transmitted diseases to them; one possible candidate is the stomach bacteria Helicobacter pylori.[247] The modern human papillomavirus variant 16A may descend from Neanderthal introgression.[248] A Neanderthal at Cueva del Sidrón, Spain, shows evidence of a gastrointestinal Enterocytozoon bieneusi infection.[249] The leg bones of the French La Ferrassie 1 feature lesions that are consistent with periostitis—inflammation of the tissue enveloping the bone—likely a result of hypertrophic osteoarthropathy, which is primarily caused by a chest infection or lung cancer.[250] Neanderthals had a lower cavity rate than modern humans, despite some populations consuming typically cavity-causing foods in great quantity, which could indicate a lack of cavity-causing oral bacteria, namely Streptococcus mutans.[251] Two 250,000-year-old Neanderthaloid children from Payré, France, present the earliest known cases of lead exposure of any hominin. They were exposed on two distinct occasions either by eating or drinking contaminated food or water, or inhaling lead-laced smoke from a fire. There are two lead mines within 25 km (16 mi) of the site.[252] |
解剖学 主な記事 ネアンデルタール人の解剖学 骨格 左側が人間の頭蓋骨で、右側がネアンデルタール人の頭蓋骨を復元したもの。脳梁の形状の違い(ネアンデルタール人の方が頭蓋骨の長さが長い)、より短い額 の比率、よりはっきりした眉尾根、より大きな鼻骨の突出、ピンと張った頬骨の角度、まっすぐな角度の顎、後頭部の饅頭が強調されている。 クリーブランド自然史博物館所蔵の現代ユーラシア人男性(左)とネアンデルタール人(右)の頭蓋骨復元像の比較。 ネアンデルタール人の頭蓋骨復元例の正面図と側面図。大きな円形の眼窩、まっすぐに伸びた顎、突き出た鼻梁、大きな眉尾根、後退した額、長い頭頂部の脳梁、後頭部の饅頭、窩、第3大臼歯の後ろの大きな隙間が強調されている。 ネアンデルタール人の頭蓋骨の特徴 ネアンデルタール人は典型的な現生人類よりもがっしりとした体格をしており[75]、胸郭が広く樽型であり、骨盤が広く[30][184]、前腕と前脚が比例して短かった[71][185]。 男性14人、女性7人のネアンデルタール人の長骨45本に基づくと、平均身長は男性164~168cm、女性152~156cmであった[75]。 2cm(5フィート9.4インチ)、162.9cm(5フィート4.1インチ)であったが、男性21人、女性15人に基づくと、時代の終わりに近づくにつ れて10cm(4インチ)ずつ減少している[186]。また、1900年の平均はそれぞれ163cm(5フィート4インチ)、152.7cm(5フィート 0インチ)であった[187]。 [187] 化石の記録によれば、ネアンデルタール人の成人の身長は約147.5cmから177cm(4フィート10インチから5フィート10インチ)であったが、中 にはもっと背が高くなった者もいたようである(足跡の長さに基づくと73.8cmから184.8cm、足跡の幅に基づくと65.8cmから 189.3cm)。 [188]ネアンデルタール人の体重については、26の標本から、男性では平均77.6kg(171ポンド)、女性では平均66.4kg(146ポンド) であった[189]。76kg(168ポンド)を用いて、ネアンデルタール人男性の肥満度指数は26.9-28.2と計算された。これは非常に頑健な体格 を示している[75]。脂肪の蓄積と体熱産生に関係するネアンデルタール人のLEPR遺伝子は、ウーリーマンモスの遺伝子と類似しているため、寒冷な気候 への適応であった可能性が高い[72]。 トンゲレンのガロ・ロマン博物館に描かれたネアンデルタール人の狩猟者たち ネアンデルタール人の首の椎骨は、(ほとんどの)現生人類よりも前方から後方、そして横方向に太く、おそらく異なる頭の形と大きさに対応するために安定性 をもたらしている。 [190]ネアンデルタール人の胸郭(胸郭のある部分)は現生人類とほぼ同じ大きさであったが、肋骨が長く直線的であったため、胸郭の中央から下部が広が り、胸郭下部での呼吸が強かったと考えられる。 [184][191][192]ケバラ2世の肺活量は9.04L(2.39US gal)であったと推定されているが、人間の平均的な肺活量は男性で6L(1.6US gal)、女性で4.7L(1.2US gal)である。また、ネアンデルタール人の胸部はより顕著であった(前後方向に広がっていた)。仙骨(骨盤と背骨をつなぐ部分)はより垂直に傾いてお り、骨盤に対して低い位置にあったため、背骨はあまり湾曲しておらず(前弯が少ない)、やや内側に折れ曲がっていた(内反していた)。現代の集団では、こ のような状態は人口のごく一部に見られ、腰椎化した仙骨として知られている[193]。背骨にこのような修正が加えられることで、側方(縦側方)への屈曲 が強化され、より広い胸郭下部をよりよく支えることになる。この特徴は、熱帯適応したホモ・エルガスターやホモ・エレクトスであっても、すべてのホモに とって正常なものであり、ほとんどの現生人類において胸郭が狭いという状態は、独特の特徴であると主張する者もいる[184]。 身体のプロポーションは通常、寒冷地での適応として「超極寒型」であることが挙げられているが、それは寒冷地で発達したヒトの集団のプロポーションに似て いるからであり[194]、ネアンデルタール人の体格は現生人類の中ではイヌイットやシベリアのユピックの体格に最も似ている[195]。 [185][194][196]にもかかわらず、イベリア半島など、より温暖な気候のネアンデルタール人は、依然として「超極寒」の体格を保持している [197]。 2019年、イギリスの人類学者ジョン・スチュワートと同僚たちは、ネアンデルタール人が寒いマンモスの草原よりも暖かい森林地帯を好んでいた証拠や、ネ アンデルタール人の速筋線維の割合が現生人類よりも高いことを示すDNA分析から、ネアンデルタール人が代わりにスプリントに適応していたと示唆した。彼 は、ネアンデルタール人の体のプロポーションと筋肉量の多さを、持久力重視の現代人の体格とは対照的な、スプリントへの適応と説明した[71]。1981 年、アメリカの古人類学者であるエリック・トリンカウスは、この代替的な説明に言及しているが、可能性は低いと考えている[185][199]。 顔 ネアンデルタール人の復元、ロンドン自然史博物館。 ネアンデルタール人は顎があまり発達しておらず、額がなだらかで、鼻が長く、広く、突き出ていた。ネアンデルタール人の頭蓋骨は一般的に、多くの現生人類 よりも細長いが、幅も広く、球形ではなく、後頭部のお団子[200]、あるいは「シニヨン」と呼ばれる頭蓋骨の後頭部の突出が特徴的であるが、これは現生 人類が持つ変異の範囲内である。これは、頭蓋底と側頭骨がより高く、より頭蓋骨の前方に向かって配置されていることと、頭蓋がより平らであることに起因す る[201]。 ネアンデルタール人の顔には、鼻下[202]と中顔面前突症があり、頬骨弓が現代人に比べて後方に位置し、上顎骨と鼻骨が比較的に前方に位置していること が特徴である[203]。ある研究では、これはネアンデルタール人が新皮質と社会性の発達を犠牲にして視覚能力を強化したためだと提唱した[204]。し かしこの研究は、眼球の大きさはネアンデルタール人や現生人類の認知能力を示す証拠にはならないと結論づけた他の研究者によって否定された[205]。 投影されたネアンデルタール人の鼻と副鼻腔は、一般的に肺に入るときに空気を暖め、水分を保持したと説明されている(「鼻のラジエーター」仮説) [206]。もし彼らの鼻が広かったとすれば、寒冷適応生物における一般的な狭い形状とは異なり、遺伝的なドリフトによって引き起こされたことになる。ま た、広く復元された副鼻腔は、現代人の副鼻腔に匹敵する大きさで、著しく大きいわけではない。しかし、もし副鼻腔の大きさが冷たい空気を呼吸するために重 要な要素ではないのであれば、実際の機能は不明であるため、そのような鼻を進化させるための進化的圧力の良い指標にはならないかもしれない[207]。さ らに、ネアンデルタール人の鼻と予測される軟部組織のパターンのコンピューター復元は、現代の北極圏の人々の鼻といくつかの類似性を示しており、潜在的に は、両集団の鼻が冷たく乾燥した空気を呼吸するために収束的に進化したことを意味している[73]。 ネアンデルタール人はかなり大きな顎を特徴としており、これはかつて、ネアンデルタール人の前歯の激しい摩耗によって証明された大きな咬合力への対応とし て引用された(「前歯部荷重」仮説)が、同様の摩耗傾向は現代人にも見られる。また、奥歯に比べて前歯の摩耗が多いのは、おそらく繰り返し使用することに 起因している。ネアンデルタール人の歯の摩耗パターンは、現代のイヌイットと最もよく似ている[206]。切歯は大きく、シャベル型をしており、現代人と 比べて、歯髄(歯の芯)が肥大して臼歯がかさばるタウロドンティズムの頻度が異常に高かった。タウロドンティズムは、かつてはネアンデルタール人の特徴的 な特徴であり、機械的な利点をもたらすか、反復的な使用に由来すると考えられていたが、単に遺伝的なドリフトの産物であった可能性が高い[209]。ネア ンデルタール人と現生人類の咬合力は、現在ではほぼ同じと考えられており[206]、現生人類の男性で約285N(64ポンド)、女性で約255N(57 ポンド)である[210]。 高齢のネアンデルタール人の復元 脳 ネアンデルタール人の脳容積は男性で平均1,640 cm3 (100 cu in)、女性で平均1,460 cm3 (89 cu in)[77][78]であり、現生人類の全グループの平均[79]よりもかなり大きい。 1 cu in)、女性は1,201 cm3(73.3 cu in)であった[211]。19万年前から2万5,000年前までの28の現生人類の標本では、平均は約1,478 cm3(90. 2立方インチ)であり、現生人類の脳の大きさは後期旧石器時代以降減少していることが示唆されている[212]。ネアンデルタール人の最大の脳であるア ムード1号は1,736cm3(105.9立方インチ)と算出されており、ヒト科の動物で記録された中で最大級のものである[78]。ネアンデルタール人 とヒトの幼児はともに約400cm3(24立方インチ)である[213]。 後方から見ると、ネアンデルタール人の脳梁は解剖学的に現代人よりも低く、広く、丸みを帯びている。この特徴的な形状は「en bombe(爆弾のような)」と呼ばれ、ネアンデルタール人特有のものである。ネアンデルタール人の脳は、比較的小さな頭頂葉[85]と大きな小脳によっ て特徴づけられていた。 [85][218]ネアンデルタール人の脳は後頭葉も大きく(ネアンデルタール人の頭蓋骨の解剖学的特徴である後頭部の饅頭と、彼らの頭蓋骨の幅の大きさ に関連している)、これはホモ・サピエンスと比較した脳の内部領域の比率の内的な違いを示唆しており、化石の頭蓋骨で得られた外的な測定値と一致してい る。 [204][219]彼らの脳はまた、側頭葉の極が大きく[218]、眼窩前頭皮質が広く[220]、嗅球が大きい[221]ことから、言語理解や感情と の関連付け(側頭葉機能)、意思決定(眼窩前頭皮質)、嗅覚(嗅球)における違いの可能性が示唆されている。彼らの脳はまた、脳の成長と発達の速度の違い も示している[222]。このような違いはわずかではあるが、自然淘汰の目に見えるものであっただろうし、社会的行動、技術革新、芸術的成果などの物質的 記録の違いの根底にあり、それを説明するものかもしれない[18][223]。 髪と肌の色 ネアンデルタール人においては、日光の不足が明るい肌の普及につながった可能性が高い[224]が、現代のヨーロッパ人における明るい肌は、おそらく青銅 器時代までは特に普及しなかったと最近主張されている[225]。 [225]遺伝学的には、ネアンデルタール人にはBNC2が存在し、これは明るい肌の色と関連しているが、BNC2の第二の変異も存在し、これは現代の集 団では暗い肌の色と関連していることがUKバイオバンクで報告されている[224]。 現代人の肌や髪の色は、MC1R遺伝子によってコードされるメラノサイト刺激ホルモン(ユーメラニン(黒色色素)とフェオメラニン(赤色色素)の割合を増 加させる)によって調節されている。現生人類には、この遺伝子の機能喪失を引き起こし、色白の肌や髪の色に関連する5つの変異体が知られており、ネアンデ ルタール人には、色白の肌や赤毛に関連する可能性のあるもう1つの未知の変異体(R307G変異体)が存在する。R307G変異体は、イタリアのモン ティ・レッシーニと、おそらくスペインのクエバ・デル・シドロンで発見されたネアンデルタール人において同定された[228]。しかし、この変異体は他の 多くのネアンデルタール人において確認されていないことから、現代人と同様に、赤はおそらくそれほど一般的な髪の色ではなかったと考えられる[224]。 代謝 しかし、歯と歯のエナメル質の成長率に基づき、ネアンデルタール人は現代人よりも早く成熟したという仮説が立てられている[229][230]。 [25]成熟における主な違いは、ネアンデルタール人の方が現代人よりも首のアトラス骨と中胸椎の癒合が約2年遅いことであるが、これは成長速度というよ りも解剖学的構造の違いによるものである可能性が高い[231][232]。 一般的に、ネアンデルタール人のカロリー必要量に関するモデルは、ネアンデルタール人が筋肉量が多く、成長速度が速く、寒さに対する体熱産生が大きいため に基礎代謝量(BMR)が高く、[233][234][235]、採食中の毎日の移動距離が長いために1日の身体活動レベル(PAL)が高かったと一般的 に仮定しているため、現代人の摂取量よりもかなり高い摂取量を報告している[233][234][235]。 [234][235]しかし、アメリカの考古学者ブライアン・ホケットは、高いBMRとPALを用いて、妊娠中のネアンデルタール人は1日あたり 5,500カロリーを消費していたと推定しており、これは大型獣肉に大きく依存する必要があったことを意味する。 ネアンデルタール人は、冬に昼間の時間が短くなる地域に住んでいたこと、大型の獲物を狩猟していたこと(このような捕食者は待ち伏せ戦術を強化するために 通常夜間に狩猟を行う)、大きな目と視覚処理神経中枢を持っていたことから、白昼よりもむしろ薄暗い時間帯に活動的であった可能性がある。クロアチアの ヴィンディヤ洞窟で発見されたネアンデルタール人は、色覚に影響を与える可能性のあるオプシン遺伝子の置換を持っていた。しかし、これらの置換の機能的な 意味については結論が出ていない[236]。ASB1とEXOC6に近いネアンデルタール人由来の対立遺伝子は、夜型人間、ナルコレプシー、昼間の昼寝と 関連している[224]。 病理学 ネアンデルタール人は高い確率で外傷を負っており、標本の推定79-94%が治癒した大きな外傷の証拠を示しており、そのうちの37-52%が重傷を負っ ており、13-19%が成人になる前に負傷していた。 [237] 極端な例として、シャニダール1号が挙げられる。シャニダール1号は、思春期に骨折した後の非結合によるものと思われる右腕の切断、左鎖骨の骨髄炎(骨の 感染症)、異常な歩行、左目の視力障害、難聴[238](おそらく水泳耳)の可能性を示している。 [239]1995年にトリンカウスは、約80%が40歳になる前に負傷に屈して死亡したと推定し、ネアンデルタール人が危険な狩猟戦略をとっていたと理 論化した(「ロデオライダー」仮説)。 [しかし、ネアンデルタール人と中期旧石器時代の現生人類の間で頭蓋外傷の割合に有意な差はなく(ネアンデルタール人の方が死亡リスクが高かったようであ るが)[240]、40歳以降に死亡した後期旧石器時代の現生人類とネアンデルタール人の標本はほとんどなく[182]、両者の間には全体的に類似した傷 害パターンが見られる。2012年にトリンカウスは、ネアンデルタール人はその代わりに、対人暴力など現代人と同じような方法で自らを傷つけたと結論づけ た[241]。ネアンデルタール人の標本124体を調査した2016年の研究では、高い外傷率はその代わりに動物による攻撃によって引き起こされたと主張 し、標本の約36%がクマによる攻撃、21%が大型ネコによる攻撃、17%がオオカミによる攻撃の犠牲者であることを発見した(合計92の陽性例、 74%)。ハイエナに襲われたケースはなかったが、それでもハイエナは少なくとも日和見的にネアンデルタール人を襲ったのであろう[242]。このような 激しい捕食は、おそらく食料と洞窟のスペースをめぐる競争による一般的な対立と、ネアンデルタール人がこれらの肉食動物を狩ったことに起因している [242]。 黒い背景にほぼ完全な骨格が水平に並べられている。 パリの人間博物館にあるラ・フェラッシー1号 人口が少なかったために遺伝的多様性が低く、おそらく近親交配が行われ、有害な突然変異をフィルタリングする集団の能力が低下した(同系交配抑制)。しか し、このことがネアンデルタール人一人の遺伝的負担にどのような影響を与えたのか、またそのために現代人よりも高い確率で先天性欠損症が発生したのかどう かは不明である。 [244] おそらく高齢(60代か70代)のためと思われるが、ラ・シャペル・オー・サント1号には脊椎に影響を及ぼすバーストルプ病と変形性関節症の徴候があった [245] 。30歳か40歳くらいで死んだと思われるシャニダール1号は、動きを制限する可能性のある変性疾患であるびまん性特発性骨格過骨症(DISH)の最も古 い症例と診断された。 ネアンデルタール人はいくつかの感染症や寄生虫に罹患していた。ヘリコバクター・ピロリという胃の細菌がその候補のひとつである[247]。現代のヒト・ パピローマ・ウイルスのバリアント16Aはネアンデルタール人の侵入から派生した可能性がある[248]。スペインのクエバ・デル・シドロンで発見された ネアンデルタール人には胃腸のエンテロシトゾーン・ビエネウシに感染した形跡がある。 [249] フランスのLa Ferrassie 1の脚の骨には、骨膜炎-骨を包む組織の炎症-に一致する病変が見られる。250] ネアンデルタール人は、典型的な虫歯の原因となる食物を大量に摂取している集団があるにもかかわらず、現代人よりも虫歯の発生率が低かった。 フランスのペイレで発見された25万年前のネアンデルタール人の2人の子供は、ヒト科の動物で鉛に暴露された最古の事例である。彼らは、汚染された食物や 水を飲んだり食べたりするか、火事の際に鉛の混じった煙を吸い込むことによって、2回にわたって暴露された。遺跡から25km(16マイル)以内には2つ の鉛鉱山がある[252]。 |
![]()

Neanderthal composite skeleton at the American Museum of Natural History
+++
Links
リンク
文献
その他の情報