[W]e
cannot factor a complex social situation into so much biology on one
side, and so much culture on the other. We must seek to understand the
emergent and irreducible properties arising from an inextricable
interpenetration of genes and environment. - Stephen Jay Gould, An Urchin in the Storm (1987) = 複雑な社会状況を、一方では生物学的に、他方では文化的にと考えることはできない。私たちは、遺伝子と環境の不可分な相互浸透から生じる、創発的で不可逆的な特性を理解しようとしなければならない。
同じ遺伝的性質をもつ集団を、異なる環境におくと、 後天的 に同じ影響をうけるはずが、多様な適応形態をもつようになる。これを後成=エピジェネシスと呼ぶ。
エビジェネシス現象は、環境や成長や加齢な どの影響により潜在的に隠れていた遺伝的形質が発現することである。育ち(そだち) Nurtureという現象は、その氏(うじ) Natureの性質を抜きに理解することはできない。氏(うじ) による性質(Nature)は、育ち(Nurture)の関係でソフトに変化することを忘れてはならない、という教訓をここで轢き出すことができる。
エピジェネシスを研究する学問を、エピジェネティクス(epigenetics)という。これまで遺伝形質はDNAに支配されている(→「セントラルドグマ」)と言われてきた。。だがDNAにはヒストンというタンパク質が複雑に階層構造化して、クロマチンや染色体の構造を形づくる。当然、それらがDNAの発現のメカニズムに関わっている可能性もある。エピジェネシス現象は、そのために、DNAだけによらない遺伝の仕組みということもできる。そして、エピジェネティクスは、DNAだけでは決まらない遺伝学であるということもできる(太田 2013)。つまり、「DNAの塩基配列を変えずに細胞が遺伝子の働きを制御する仕組みを研究する学問(研究)」をエピジェネティクスと呼ぶ。
エピジェネティクス(英
語:
epigenetics)、後成学(こうせいがく)または後成遺伝学(こうせいいでんがく)とは、一般的には「DNA塩基配列の変化を伴わない細胞分裂後
も継承される遺伝子発現あるいは細胞表現型の変化を研究する学問領域」である[1][2]。ただし、歴史的な用法や研究者による定義の違いもあり、その内
容は必ずしも一致したものではない[3]。特に遺伝子(gene)ではなくゲノム(genome)を対象とする場合、エピゲノミクスあるいはエピゲノムと
呼ばれることもある。
多くの生命現象に関連し、人工多能性幹細胞(iPS細胞)・胚性幹細胞(ES細胞)が多様な器官となる能力(分化能)、哺乳類クローン作成の成否と異常発
生などに影響する要因(リプログラミング)、がんや遺伝子疾患の発生のメカニズム、脳機能[4][5]などにもかかわっている。
遺伝形質の発現は、セントラルドグマ説[6][7]で提唱されたようにDNA複製→RNA転 写→タンパク質への翻訳→形質発現の経路により、DNAに記録されている遺伝情報が表現型として実現した結果とされてきた。セントラルドグマにおける形質 の変化とは、遺伝情報の変化であり、その記録媒体であるDNA塩基配列の変化が原因となっている。レトロウイルスやレトロトランスポゾンによるRNAから DNAへの情報の還元という例外を含みながらも、従来の分子生物学・遺伝学ではセントラルドグマを基礎においた研究が行われてきた[8]。
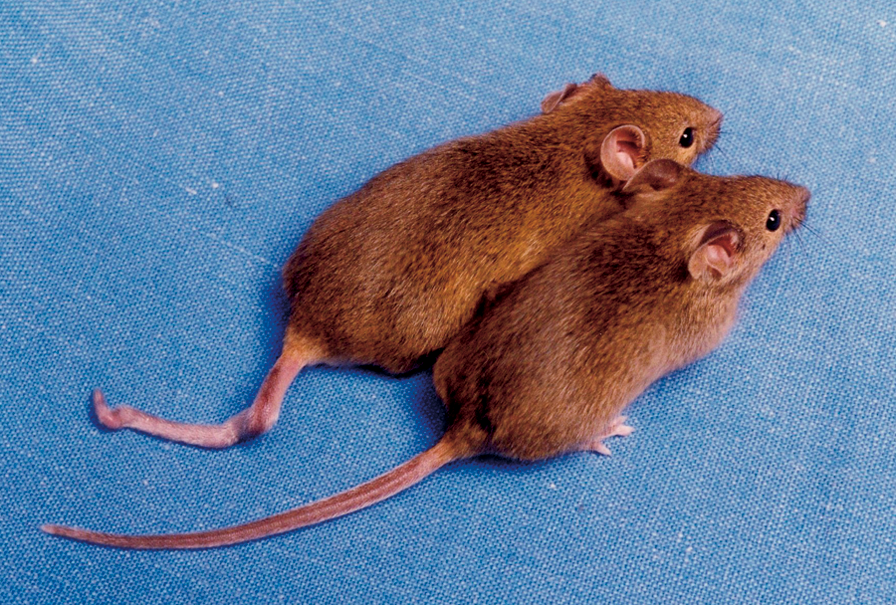
DNAメチル化の差によって尾の形状が異なる二匹のクローンマウス[9]
しかしながら、先天的には同じ遺伝情報、つまり同じゲノム(DNA塩基配列)であっても、細胞レベルあるいは個体レベルの形質の表現型が異なる例もまれではない。
たとえば動物では、単細胞である受精卵から発生し、胚の全能性幹細胞はさまざまな多能性細胞系列となり、さらに器官ごとに異なった細胞に分化し、それぞれ
の器官・細胞は異なる機能を分担している。この過程で細胞は、分化の経歴と存在する部位に依存して、ある遺伝子を抑制する一方で、他のある遺伝子は活性化
している[10]。また一卵性双生児やクローン動物、あるいは挿し木や球根・地下茎などの栄養生殖で増殖した植物でも、遺伝子型は同一にもかかわらず個体
間に違いが認められることが多い。
このような例は、細胞レベルではシグナル伝達による細胞間の応答反応、個体レベルでは環境と遺伝の相互作用によって主に説明がなされていた。しかしなが
ら、細胞がどのように経歴を「記憶」するのか、個体間の表現型の差がどのように生じるかは、遺伝子機能の面からは明らかにされていない部分があった。
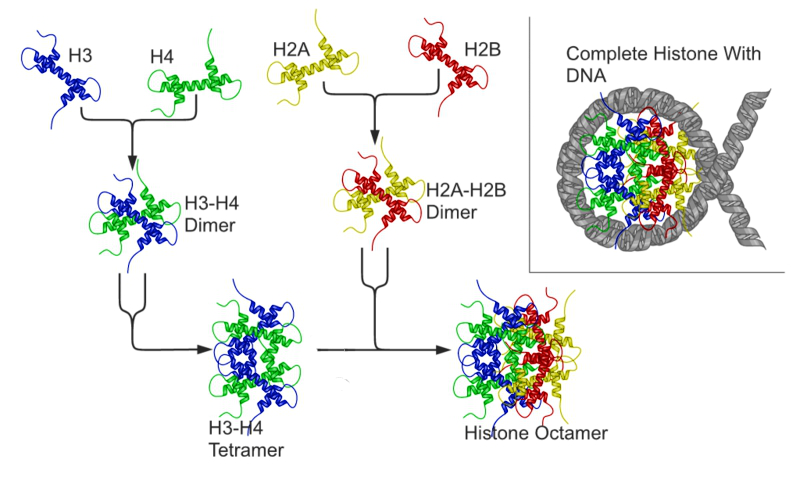
クロマチン中のヒストンとDNAの模式図
(右上)ヌクレオソーム構造: ヒストン八量体に巻き付いたDNA
(上段) H3, H4, H2A, H2B コアヒストン単量体(モノマー)
(中・下段) H3-H4テトラマーとH2A-H2Bダイマー 2個が会合し、ヒストン八量体となる。
1942年にコンラッド・H・ウォディントン(英語版)は、「遺伝物質からはじまり最終的な
生物を形づくるすべての制御された過程」言い換えると「遺伝子が表現型を作るために周辺環境とどのように相互作用するのか」を表現するために、「エピジェ
ネティクス」という用語を作成した[2][11]。その後、エピジェネティクスは、DNA塩基配列の変化を伴わない後天的な遺伝子制御の変化を主な対象と
した研究分野となり、各種生物のゲノムの解読が進んだ2000年代以降、エピジェネティクス研究が盛んになってきている。
前述の通り「エピジェネティクス」の内容は普遍的に定義されたものではない[3]。しかしながら、入門的な解説の場合、表1に示す各種の過程のうち染色体クロマチンを構成するDNAのメチル化およびヒストンの化学的修飾に重点を置いて説明される[12]。
この場合、DNA塩基配列の変化つまり突然変異と、エピジェネティック(=エピジェネティクス的)制御とは独立である。それらは、同一個体内での組織の違
いあるいは個体発生・細胞分化の時間軸上の違いで生じる変化である。しかしそれらと異なり、変化した表現型が個体の世代を超えて受け継がれる「エピジェネ
ティック遺伝」の例も見出されており、研究が進められている[13][14]。これは、ある生物におけるエピジェネティックな変化がそのDNAの基本構造
を変えることができるかどうかというラマルキズム型の問題を提起する。

「分裂して生じる2つの娘細胞は遺伝子の変化に関し
て同等ではない。分裂後、ある細胞では特定の遺伝子が活性化されるだろうし、別の遺伝子はそこに有りながら不活性化される。このような活性化や不活性化
は、遺伝子がクロマチン物質によって覆われているが故に生じる。遺伝子の活性化は、覆われていた遺伝子が遺伝子が露出したときにのみ起こるだろう」——マ
クリントック(1951)、ただし翻訳引用は(太田 2013:89)。
★用語集
| ヒストン | histone | 真核生物のクロマチン(染色体)を構成する主要なタンパク質 |
| 転移性因子 | トランスポゾンやレトロトランスポゾンなどからなる(太田 2013:81) | |
| ヒストン修復 | ||
| メチル化 | ||
| メタ情報 | ||
| DNA脱メチル化酵素 | ||
| 飢餓ストレス | ||
| エピゲノム |
ゲ
ノムDNA中の遺伝子をRNAに転写する度合いを制御する、ゲノムDNAの可逆的な化学修飾の仕組みをいう。細胞核に収められたゲノムDNAはクロマチン
[4]と呼ばれる凝縮した構造をとっており、エピゲノムは転写前にその凝縮を解きほぐすなど、クロマチン構造を変換する複数の反応素過程を制御する。https://www.riken.jp/press/2020/20201126_1/index.html |
|
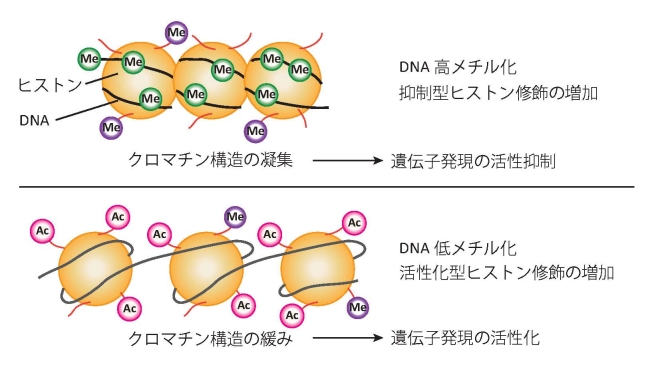
|
図3:遺伝子発現調節に関わるエピジェネティック修飾変化の模式図(→国立環境研究所「エピジェネティクス」) 上:DNA の高メチル化や抑制型ヒストン修飾の増加によりクロマチン構造が凝集すると、遺伝子発現の活性が抑制されます。 下:DNA の低メチル化や活性化型ヒストン修飾の増加によりクロマチン構造が緩むと遺伝子発現が活性化されます。 |
|
リンク
- 社会生物学/包括適応度/ハーツォグ・ノート/////////
- ローカル・バイオロジーズ
- 認知症・経済格 差・社会関係資本・トラウマ -レジリアンス
- 民俗学とプラグマティックな医学について
- 医療人類学辞典
- 太田邦史研究室
文献
- エピゲノムと生命 : DNAだけでない「遺伝」のしくみ / 太田邦史著、講談社 , 2013年
- ジャンクDNA : ヒトゲノムの98%はガラクタなのか? / ネッサ・キャリー著 ; 中山潤一訳、丸善出版 , 2016年
- やわらかな遺伝子 / マット・リドレー著 ; 中村桂子, 斉藤隆央訳, 紀伊國屋書店 , 2004年/Nature via nurture : genes, experience and what makes us human, M. Ridley,
- Margaret Lock, 2013. The lure of the epigenome. Lancet Volume 381, Issue 9881, 1–7 June 2013, Pages 1896-1897. https://doi.org/10.1016/S0140-6736(13)61149-6
その他の情報


