縄文人
Jōmon people
☆ 縄文人(じょうもんじん)とは、縄文時代(新石器時代)に日本列島全域に居住してい た人々の総称。約1万6000年前から約3000年前まで(→縄文時代)現在の北海 道から沖縄本島にかけて住み、縄文文化と呼ばれる文化形式を保持していた。※「縄 文人」は独自進化したアジアの特異集団だった 読売新聞、2017年12月15日.
★海部陽介によると、ホモ・サピエンスが日本列島にやってきたのは、今からおよそ、3万8千年前といわれ、後期旧石器文化をかたちづくる(海部 2016:118)。海部によると縄文文化の幕開けは1万6000年前ごろといわれる。また海部は、縄文人は単一の集団であったという見解は間違いであることを主張している(海部 2016:201)
★ミ
トコンドリアDNAの解析から、篠田謙一は「縄文人は旧石器時代にさかのぼる周辺の南北双方の地域から流入した人びとが、列島の内部で混合することによっ
て誕生した」と主張し、人骨形態学の片山一道も、列島内で縄文人になったとの主張を紹介している(篠田 2015:191)
★
英語のウィキペディアのエントリー:「縄文人(縄文人、縄文時代人)は、縄文時代(紀元前1万4000[〜6000]年頃~紀元前300年頃)に日本列島
に生息していた狩猟採集民の総称である。彼らは共通の縄文文化によって結びついており、その文化は定住性と文化的な複雑性の面でかなりの程度に達してい
た。縄文人は、東アジアで大きく分岐した祖先を持つという特徴があり、現代の日本人に10~20%の祖先を残した。複数の縄文時代の遺跡から得られた集団
ゲノムデータによると、縄文人は
3万8000年から2万5000年前のいつかの時点で、北方および南方の東アジア人の分岐よりも前に、「祖先的東アジア人」から分岐した。ただし、「基本
的な東アジア人」である天元人とホアビン人の系統(約3万9000年前)からは分岐した後であった。日本列島への移住後(3万8000年前?)、彼らは紀元前2万~2万5000
年頃には、外部からの遺伝子流入からほぼ孤立した状態となった。」
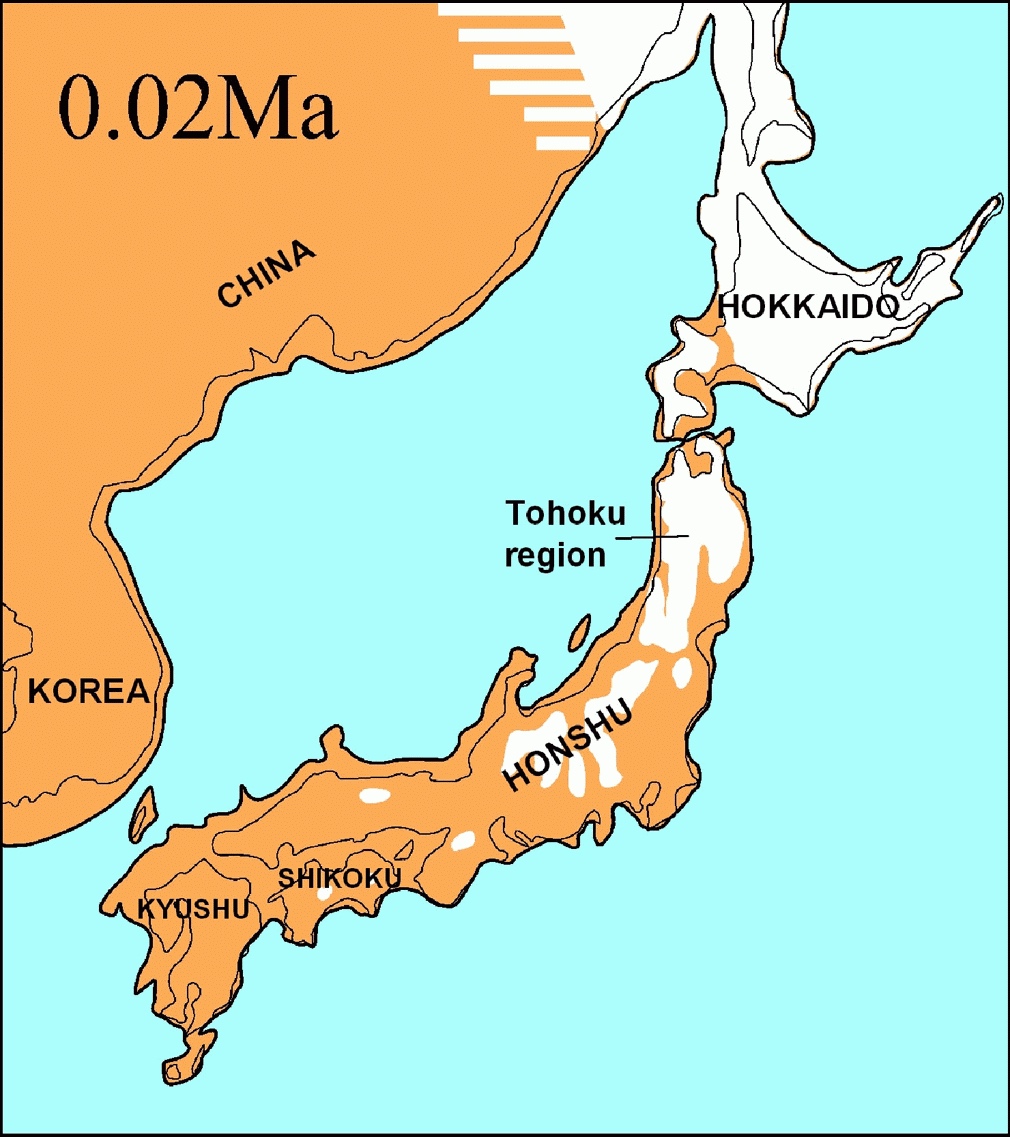

更新世の終わり2万年前頃には、ほぼ現在に近い地形であるが、最終氷期最盛期のため海面が低下し日本海と外洋を繋ぐ海峡は非常に狭かった。(0.02Ma)(「日本列島」より)/ Diorama of Jomon people at Sannai Maruyama.(三内丸山遺跡)
| Jōmon people
(縄文 人, Jōmon jin) is the generic name of the indigenous hunter-gatherer
population that lived in the Japanese archipelago during the Jōmon
period (c. 14,000 to 300 BC). They were united through a common Jōmon
culture, which reached a considerable degree of sedentism and cultural
complexity. The Jōmon people are characterized by a deeply diverged East Asian ancestry and contributed around 10–20% ancestry to modern Japanese people.[1][2][3][4] Population genomic data from multiple Jōmon period remains suggest that they diverged from "Ancestral East Asians" prior to the divergence of Northern and Southern East Asians, sometime between 38,000 and 25,000 years ago, but after the divergence of "Basal East Asian" Tianyuan and Hoabinhian lineages (c. 39,000 years ago). After their migration into the Japanese archipelago, they became largely isolated from outside geneflow at c. 20,000 to 25,000 BC.[4][2][3][5] |
縄文人(縄文人、縄文時代人)は、縄文時代(紀元前1万4000年頃~紀元前300年頃)に日本列島に生息していた狩猟採集民の総称である。彼らは共通の縄文文化によって結びついており、その文化は定住性と文化的な複雑性の面でかなりの程度に達していた。 縄文人は、東アジアで大きく分岐した祖先を持つという特徴があり、現代の日本人に10~20%の祖先を残した。[1][2][3][4] 複数の縄文時代の遺跡から得られた集団ゲノムデータによると、縄文人は 3万8000年から2万5000年前のいつかの時点で、北方および南方の東アジア人の分岐よりも前に、「祖先的東アジア人」から分岐した。ただし、「基本 的な東アジア人」である天元人とホアビン人の系統(約3万9000年前)からは分岐した後であった。日本列島への移住後、彼らは紀元前2万~2万5000 年頃には、外部からの遺伝子流入からほぼ孤立した状態となった。[4][2][3][5] |
| Culture See also: Jōmon period The culture of the Jōmon people was largely based on food collection and hunting, but it is also suggested that the Jōmon people practiced early agriculture.[6] They gathered tree nuts and shellfish, were involved in hunting and fishing, and also practiced some degree of agriculture, such as the cultivation of the adzuki bean and soybean.[7] The Jōmon people also used stoneware and pottery, and generally lived in pit dwellings.[8] Some elements of modern Japanese culture may have come from the Jōmon culture. Among these elements are the precursory beliefs to modern Shinto, some marriage customs, some architectural styles, and possibly some technological developments such as lacquerware, laminated yumi, metalworking, and glass making. Pottery The style of pottery created by the Jōmon people is identifiable for its "cord-marked" patterns, hence the name "Jōmon" (縄文, "straw rope pattern"). The pottery styles characteristic of the first phases of Jōmon culture used decoration created by impressing cords into the surface of wet clay, and are generally accepted to be among the oldest forms of pottery in East Asia and the world.[9] Next to clay pots and vessels, the Jōmon also made many highly stylized statues (dogū), clay masks, stone batons or rods and swords.[10] Craftsmanship  Magatama – kidney-shaped beads – are commonly found in Jōmon period Japanese finds, as well as in parts of Northeast Asia and Siberia. There is evidence that the Jōmon people built ships out of large trees and used them for fishing and traveling; however, there is no agreement as to whether they used sails or paddles.[11] The Jōmon people also used obsidian, jade and different kinds of wood.[12] The Jōmon people created many jewelry and ornamental items; for instance, magatama were likely invented by one of the Jōmon tribes, and are commonly found throughout Japan and less in Northeast Asia.[10] Religion Main article: Ko-Shintō It is suggested that the religion of the Jōmon people was similar to early Shinto (specifically Ko-Shintō). It was largely based on animism, and possibly shamanism. Other similar religions are the Ryukyuan and Ainu religions.[13] |
文化 関連項目:縄文時代 縄文人の文化は主に食料の採集と狩猟を基盤としていたが、初期の農業も行っていた可能性がある。[6] 彼らは木の実や貝類を集め、狩猟や漁労に従事し、小豆や大豆などの栽培といったある程度の農業も行っていた。[7] 縄文人は土器や陶器も使用し、一般的に竪穴住居に住んでいた。[8] 現代の日本文化のいくつかの要素は、縄文文化に由来する可能性がある。その要素には、現代の神道の先行する信仰、一部の結婚の風習、一部の建築様式、そして漆器、積層弓、金属加工、ガラス製造などの技術開発が含まれる可能性がある。 土器 縄文人が作った土器のスタイルは、「縄目」模様で識別でき、それが「縄文」(縄目文様)という名称の由来となっている。縄文文化の初期段階に特徴的な土器 の様式は、濡れた粘土の表面に縄を押し当てて装飾を施したもので、一般的に東アジアおよび世界で最も古い土器の形式のひとつであると考えられている。 [9] 縄文人は、土器や容器のほかにも、高度に様式化された多くの像(土偶)、粘土の仮面、石の棒や剣も製作した。[10] 職人技 腎臓形の勾玉(まがたま)は、縄文時代の日本の遺物として一般的に見つかるもので、北東アジアやシベリアの一部でも見つかっている。 縄文人は大きな木から船を造り、漁や移動にそれを使っていたという証拠があるが、帆を使ったのか、櫂を使ったのかについては意見が一致していない。 [11] 縄文人は黒曜石や翡翠 また、さまざまな種類の木材も使用していた。[12] 縄文人は多くの装飾品や装飾品を作った。例えば、勾玉は縄文人の部族の1つによって発明された可能性が高く、日本全土で一般的に見つかっており、北東アジ アではあまり見つかっていない。[10] 宗教 詳細は「古神道」を参照 縄文人の宗教は初期の神道(特に古神道)に類似していたと推測されている。それは主に精霊説を基盤としており、おそらくはシャーマニズムも含まれていた。類似した宗教としては琉球神道やアイヌ神道がある。[13] |
| Languages It is not known what language or languages were spoken in Japan during the Jōmon period. Suggested languages are: the Ainu language, Japonic languages, Austronesian languages, or unknown and today extinct languages.[14][15] While the most supported view is to equate the Ainu language with the Jōmon language, this view is not uncontroversial or easily acceptable as there were probably multiple distinct language families spoken by the Jōmon period population of the Japanese archipelago.[16] Alexander Vovin (1993) argues that the Ainu languages originated in Central Honshu, and were later pushed northwards into Hokkaido, where the early Ainu-speakers merged with local groups, forming the historical Ainu ethnicity. Bilingualism between Ainu and Japanese was common in Tohoku until the 10th century.[17][18] According to Vovin (2021) there is also some evidence for the presence of Austronesian languages close to the Japanese archipelago, which may have contributed some loanwords to the early Japanese.[18] Some linguists suggest that the Japonic languages may have been already present within the Japanese archipelago and coastal Korea, before the Yayoi period, and can be linked to one of the Jōmon populations of southwestern Japan, rather than the later Yayoi or Kofun period rice-agriculturalists. Japonic-speakers then expanded during the Yayoi period, by assimilating the newcomers, adopting rice-agriculture, and fusing mainland Asian technologies with local traditions.[19] |
言語 縄文時代に日本列島で話されていた言語は不明である。推定される言語は、アイヌ語、日本語、オーストロネシア語族、または不明で現在では絶滅した言語であ る。[14][15] 最も有力な説は、アイヌ語を縄文語と同一視するものであるが、この説には異論があり、また、日本列島に縄文時代に暮らしていた人々が話していたのは、おそ らく複数の異なる語族であったため、容易に受け入れられるものではない。[16] アレクサンダー・ヴォヴィン(1993年)は、アイヌ語は本州中部で発生し、後に北方に押しやられて北海道に到達し、初期のアイヌ語話者は地元のグループ と融合し、歴史的なアイヌ民族を形成したと主張している。東北地方では、10世紀までアイヌ語と日本語のバイリンガリズムが一般的であった。[17] [18] ヴォヴィン(2021年)によると、オーストロネシア語族の言語が日本列島に近い場所に存在していたことを示す証拠もいくつかあり、初期の日本語にいくつ かの借用語がもたらされた可能性がある。[18] 一部の言語学者は、ヤポニック語族は弥生時代以前にすでに日本列島と朝鮮半島沿岸部に存在しており、後の弥生時代や古墳時代の稲作農耕民ではなく、日本の 南西部の縄文人の集団のひとつと関連している可能性があると指摘している。ヤポニック語族は、弥生時代に新参者と融合し、稲作農耕を取り入れ、アジア大陸 の技術と地元の伝統を融合させることで拡大した。[19] |
| Origins See also: Genetic history of East Asians The Jōmon people represent the descendants of the Paleolithic inhabitants of the Japanese archipelago, which became isolated from other mainland Asian groups some 22,000 to 25,000 years ago. They have been described as "one of the most deeply diverged populations in East Asia".[20][21][5][22] Genetics  Phylogenetic position of the Jōmon lineage among other East Eurasians 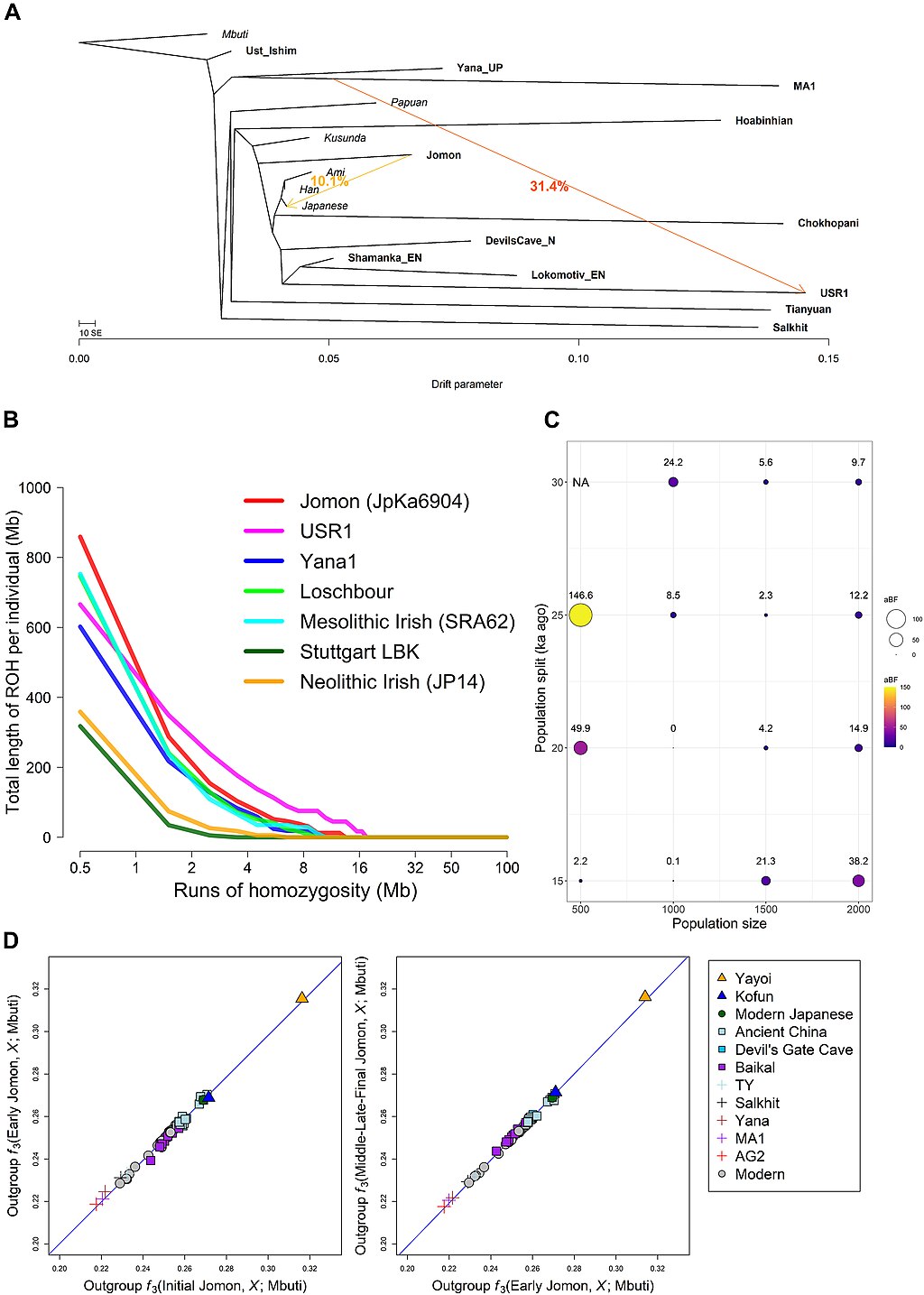 Demographic history of the Jomon lineage (A) Maximum likelihood phylogenetic tree reconstructed by TreeMix under a model of two migrations 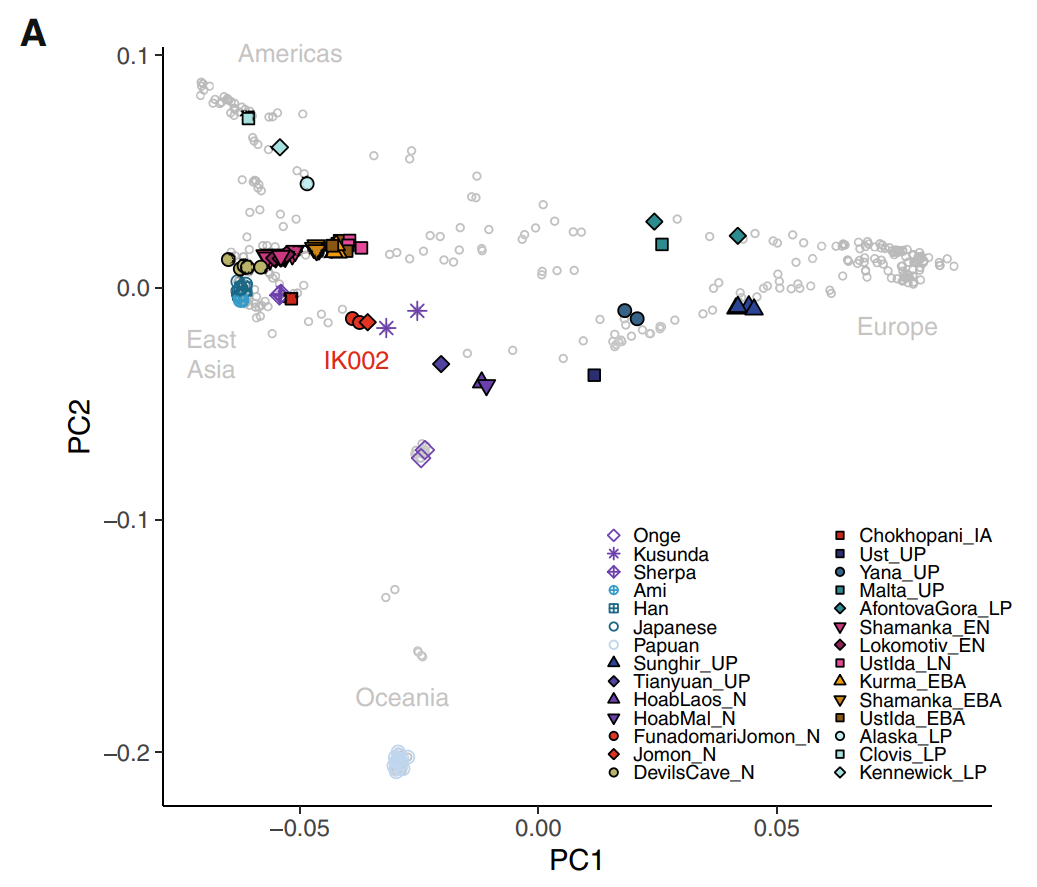 Principal component analysis (PCA) of ancient and present-day individuals from worldwide populations The Jōmon lineage is inferred to have diverged from Ancient East Asians between 25–38,000 years ago, before the divergence of Ancient Northern East Asians and Ancient Southern East Asians, but after the divergence of the Upper Paleolithic Tianyuan lineage and ancient Southeast Asian hunter-gatherers (Hoabinhians).[23][4][21] Like other East Asian populations, the ancestors of the Jōmon people originated from Southeast Asia and expanded to East Asia via a coastal route. They represent one of the "earliest waves of migration".[24][21][25][26] The Jōmon lineage displays a closer genetic affinity to Ancient Northern and Southern East Asian lineages, compared to Basal East Asian Tianyuan or Hoabinhian lineages.[21] Beyond their genetic affinity with other Eastern Asian lineages, the Jōmon also display a weak yet marginally significant genetic affinity with the Yana Rhinoceros Horn Site specimen, associated with Ancient North Eurasians (or Ancient North Siberians), which may point to geneflow between both groups prior to their isolation from other East Eurasian populations.[4] This geneflow may be associated with the introduction of microblade technology to northern Japan.[2][27] 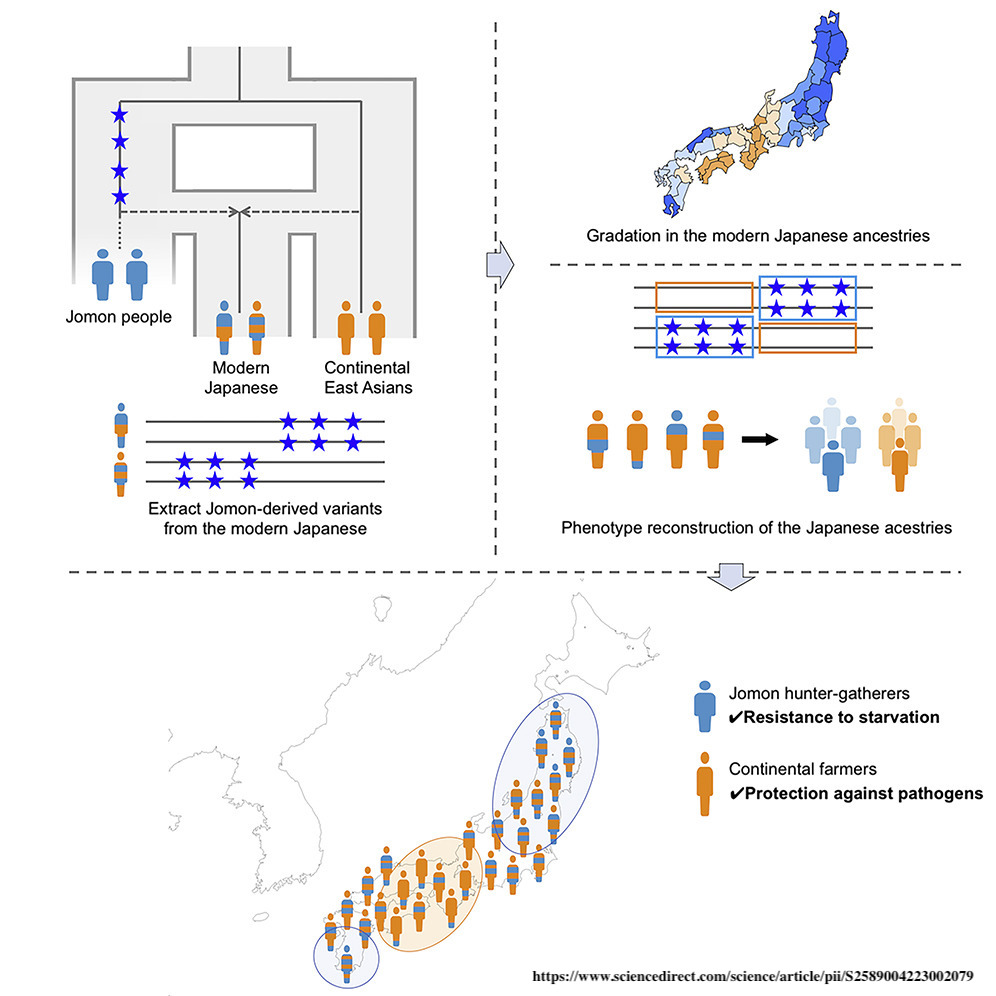 Jomon and Continental Asian contributions to modern Japanese Full genome studies on multiple Jōmon remains revealed them to carry gene alleles associated with a higher alcohol tolerance, wet earwax, no derived variant of the EDAR gene, and that they likely frequently consumed fatty sea and land animals. They also carried alleles for medium to light skin, dark and fine/thin hair, and brown eyes. Some samples also displayed a higher risk of developing liver spots if spending too much time in the sun.[25][28][2] Genetic data further indicates that the Jōmon peoples were genetically predisposed for short stature, as well as higher triglyceride and blood sugar levels, increasing the risk of obesity. At the same time, it gave them resistance to starvation. Modern Japanese share these alleles with the Jōmon period population, although at lower and variable frequency, inline with the inferred admixture among modern Japanese peoples. Watanabe et. al (2023) states that the genetic predisposition for shorter stature among Japanese people often correlates with high Jōmon ancestry, with the opposite correlating with high continental East Asian ancestry.[25] Liu et. al (2024) stated that Jōmon admixture in contemporary Japanese people varies depending on region, with admixture being the highest in southern Japan, especially Okinawa (28.5%), followed by northeastern Japan (19%) and western Japan (12%).[29] Previous studies showed that populations from Tohoku, Kanto and Kyushu were more genetically related to Ryukyuan populations whilst populations from Kinki and Shikoku were more related to continental East Asian populations.[25] Haplogroups It is thought that the haplogroups D-M55 (D1a2a) and C1a1 were frequent among the historical Jōmon period people of Japan. O-M119 is also suggested to have been presented in at least some Jōmon period remains. One 3,800 year old Jōmon man excavated from Rebun Island was found to belong to Haplogroup D1a2b1(D-CTS 220).[30] Today, haplogroup D-M55 is found in about 35%[31] and haplogroup C1a1 in about 6% of modern Japanese people. D-M55 is found regularly only in Japanese (Ainu, Ryukyuans, and Yamato) and, albeit with much lower frequency, in Koreans.[32] D-M55 also has been observed in Micronesia 5.1%, Timor 0.2%, China 0–0.4%, this is explained by recent admixture, dating back to the Japanese empire (1868-1945) occupation of those regions.[32] Haplogroup C1a1 has been found regularly in about 6% of modern Japanese. Elsewhere, it has been observed sporadically in individuals from South Korea, North Korea (South Hwanghae Province), and China (ethnic Korean in Ning'an and Han Chinese in Linghai, Guancheng Hui District, Haigang District, and Dinghai District).[33] A 2021 study estimated that the frequency of the D-M55 clade increased during the late Jōmon period.[34] The divergence between the D1a2-M55 and the D1a-F6251 subclades (the latter of which is common in Tibetans, other Tibeto-Burmese groups, and Altaians, and has a moderate distribution in the rest of East Asia, Southeast Asia, and Central Asia) may have occurred near the Tibetan Plateau.[31] The MtDNA haplogroup diversity of the Jōmon people is characterized by the presence of haplogroups M7a and N9b. Studies published in 2004 and 2007 show the combined frequency of M7a and N9b observed in modern Japanese to be from 12~15% to 17% in mainstream Japanese.[35][36] N9b is frequently found among the Hokkaido Jomons while M7a is found frequently among the Honshu Jomons.[37] However N9b is found only at very low percentage among the Honshu Jomon.[37] M7a is estimated to share a most recent common ancestor with M7b'c, a clade whose members are found mainly in Japan (including Jōmon people), other parts of East Asia, and Southeast Asia, 33,500 (95% CI 26,300 <-> 42,000) years before present.[38] All extant members of haplogroup M7a are estimated to share a most recent common ancestor 20,500 (95% CI 14,700 <-> 27,800) years before present.[38] Haplogroup M7a now has its highest frequency in Okinawa. Morphological characteristics Skull_of_Jomon_people_man.jpg Male skull of the late Jōmon period (replica). Excavated at Miyano Kaizuka (Iwate Prefecture). Exhibition in National Museum of Nature and Science.[39] Several studies of numerous Jōmon skeletal remains that were excavated from various locations in the Japanese archipelago allowed researchers to learn more about the Jōmon period population of Japan. The Jōmon people were relatively close to other East Asian people, however shared more similarities with Native American samples. Within Japan, regional variance among different Jōmon remains was detected. Historically, the Jōmon people were classified as Mongoloid.[40][41] Jomon_people_Skull_and_Restoration_model_-_Niigata_Prefectural_Museum_of_History.jpg Forensic reconstruction from a Jōmon skull, displayed at Niigata Prefectural Museum of History. Dental morphology suggests that the Jōmon had Sundadont dental structure which is more common among modern Southeast Asians and indigenous Taiwanese, and is ancestral to the Sinodont dental structure commonly found among modern Northeast Asians, suggesting that the Jōmon split from the common "Ancestral East Asians" prior to the formation of modern Northeast Asians.[42] Kondo et al. 2017, analyzed the regional morphological and craniometric characteristics of the Jōmon period population of Japan, and found that they were morphologically heterogeneous and displayed differences along a Northeast to Southwest cline. Differences were based on the cranial index, with Hokkaido Epi-Jomon crania being mesocephalic and Okinawan crania being brachycephalic. They concluded that the "Jomon skulls, especially in the neurocranium, exhibit a discernible level of northeast-to-southwest geographical cline across the Japanese archipelago, placing the Hokkaido and Okinawa samples at both extreme ends. The following scenarios can be hypothesized with caution: (a) the formation of Jomon population seemed to proceed in eastern or central Japan, not western Japan (Okinawa or Kyushu regions); (b) the Kyushu Jomon could have a small-sized and isolated population history; and (c) the population history of Hokkaido Jomon could have been deeply rooted and/or affected by long-term extrinsic gene flows." They also suggested that regional differences in cranial length is based on genetic, or phylogenetic, and environmental effects.[43] But Gakuhari et al. 2020 clarifies that Hokkaido and Honshu Jōmon cluster together and form a clade to the exclusion of other groups.[2] According to Chatters et al., the Jōmon display some similarities to the Native American Kennewick Man.[44] Chatters, citing anthropologist C. Loring Brace, classified Jōmon and Polynesians as a single craniofacial "Jōmon -Pacific" cluster.[45] Chatters, citing Powell, argues that the Jōmon most resembled the Native American Kennewick Man and Polynesians. According to him, the Ainu descend from the Jōmon people, an East Asian population with "closest biological affinity with south-east Asians rather than western Eurasian peoples".[44][46] Powell further elaborates that dental analysis showed the Jōmon to be of the Sundadont type.[45] According to Matsumura et. al (2019), the Jōmon shared similar phenotypes with prehistoric south Chinese and Southeast Asian peoples, such as 'dolichocephalic calvaria, large zygomatic bones, remarkably prominent glabellae and superciliary arches, concave nasal roots, and low and wide faces'.[47] According to Wang et. al (2021), these peoples were still related to later East Asians despite phenotypic discontinuities.[48] Craniofacial features of the Jōmon people were significantly retained by the Ainu and Okinawans/Ryukyuans.[49] The former have 2 genes "associated with facial structure in Europeans" but still possessed hair and teeth morphology found in East Asians.[50] In regards to facial flatness, the Ainu were intermediate between Caucasoids and Mongoloids[51] but another study states that they were well within the Mongoloid range.[52] The latter have a "well-defined and less flat upper face", which is characterized by a prominent glabella and nasal root.[49] ATL retrovirus A gene common in Jōmon people is a retrovirus of ATL (human T lymphotropic virus, HTVL-I). This virus was discovered as a cause of adult T cell leukemia (ATL), and research was advanced by Takuo Hinuma of Kyoto University Virus Research Institute. Although it was known that many virus carriers existed in Japan, it was not found at all in neighboring countries of East Asia. Meanwhile, it has been found in many Africans, Native Americans, Tibetans, Siberians, Burmese people, Indigenous people of New Guinea, Polynesians, etc. Looking at distribution in Japan, it is seen particularly frequently in southern Kyushu, Nagasaki Prefecture, Okinawa and among the Ainu. And it is seen at medium frequency in the southern part of Shikoku, southern part of the Kii Peninsula, the Pacific side of the Tōhoku region (Sanriku) and Oki Islands. Overall, carriers of the ATL retrovirus were found to be more common in remote areas and remote islands. When examining the well-developed areas of ATL in each region of Kyushu, Shikoku, and Tōhoku in detail, carriers are preserved at high rates in small settlements that were isolated from the surroundings and inconvenient for traffic. The path of natural infection of this virus is limited to vertical infection between women and children (most often through breastfeeding) and horizontal infection between males and females (most often from males to females through sexual intercourse).[53] Based on the above, Hinuma concluded that the high frequency area of this virus indicates that high density remains of Jōmon people.[54] Contributions to other populations Historical groups Full genome analyses of Okhotsk culture remains on Sakhalin found them to be derived from three major sources, notably Ancient Northeast Asians, Ancient Paleo-Siberians, and Jōmon people of Japan. An admixture analysis revealed them to carry c. 54% Ancient Northeast Asian, c. 22% Ancient Paleo-Siberian, and c. 24% Jōmon ancestries respectively.[55] Genetic analyses on ancient remains from the southern Korean Peninsula revealed elevated Jōmon ancestry at c. 37%, while Yayoi remains in Japan were found to carry nearly equal amounts of Jōmon ancestry (35–60%) and Ancient Northeast Asian-like ancestry (40–65%). These results suggest the presence of a Jōmon-like population on the Korean peninsula and their significant contribution to the formation of early Japonic-speakers. As such, the "agricultural transition in prehistoric Japan involved the process of assimilation, rather than replacement, with almost equal genetic contributions from the indigenous Jomon" and mainland Asian migrants of the Mumun/Yayoi period.[4][56] Modern groups  The Ainu are among the modern groups displaying the highest amounts Jōmon-derived ancestry. (Takuke YAMAMOTO, 1904-1993) Jōmon-associated ancestry is commonly found throughout the Japanese archipelago, ranging from c. 15% among modern Japanese people, to c. 30% among Ryukyuan people, and up to c. 75% among modern Ainu people, and at lower frequency among surrounding groups, such as the Nivkhs or Ulch people, but also Koreans and other coastal groups, suggesting that the Jōmon were not completely isolated from other groups.[2][28][55][57][21] Wang and Wang (2022) noted that peoples with Jōmon ancestry during the Three Kingdoms period significantly contributed to the genetic makeup of modern Koreans. But this ancestry was 'diluted' over time due to subsequent arrivals of northern Han Chinese.[56] McColl et. al (2018) states that present Southeast Asians derive ancestry from at least 4 ancient population sources. One source was the mainland Hòabìnhians, who shared ancestry with the ancient Japanese Ikawazu Jōmon, along with present-day Andamanese Önge and Malaysian Jehai.[58] However, according to Yang et. al (2020), the Jōmon individual showed affinities to several coastal Neolithic populations in Siberia and southern East Asia, with the latter clustering with present-day southern East Asians.[59] Along with modern Japanese, Ulch, and Nivkhs, present-day Taiwanese aborigines (i.e. Ami and Atayal) also showed excess allele sharing with a 2500 year old individual in mainland Japan, which was characterized by 'typical Jōmon culture'.[2] |
起源 関連情報: 東アジア人の遺伝的歴史 縄文人は、約2万2000年から2万5000年前に他のアジア本土集団から孤立した、日本列島に生息していた旧石器時代の住民の子孫である。彼らは「東アジアで最も分化した集団のひとつ」と表現されている。[20][21][5][22] 遺伝学 他の東アジア人集団における縄文人の系統発生上の位置 縄文人の系統発生上の人口統計学的歴史 (A) TreeMix による2回の移住モデルに基づく最尤法系統樹 世界中の集団に属する古代および現代の個体群の主成分分析(PCA) 縄文人の系統は、古代東アジア人から2万5000年~3万8000年前に分岐したと推定されており、これは古代北東アジア人と古代南東アジア人の分岐より も前だが、旧石器時代の天 。[23][4][21] 他の東アジアの集団と同様に、縄文人の祖先は東南アジアを起源とし、海岸沿いのルートを通って東アジアに拡散した。彼らは「最も早い時期の移住の波」のひ とつである。[24][21][25][26] 縄文人の系統は、基底的な東アジア系統である天元人やホアビン人に比べ、古代の東アジアの南北の系統とより近い遺伝的類縁関係にある。[21] 他の東アジア系統との遺伝的類縁関係に加え、縄文人の系統は、 古代北ユーラシア人(または古代北シベリア人)に関連するヤナサイのサイ角遺跡の標本との間に、弱いながらも限界的に有意な遺伝的類似性も示している。こ れは、他の東ユーラシアの集団から孤立する前の両集団間の遺伝子交流を示している可能性がある。[4] この遺伝子交流は、日本北部へのマイクロブレード技術の導入に関連している可能性がある。[2][27] 縄文人と大陸アジア人の現代日本人への影響 複数の縄文人の遺骨の全ゲノム研究により、縄文人はアルコール耐性、耳垢が湿っている、EDAR遺伝子の派生変異がない、そして海産物や陸産物の脂肪分の 多い動物を頻繁に摂取していた可能性が高いことが明らかになった。また、彼らは中程度から色白の肌、黒髪で髪質は細い、瞳の色は茶色という遺伝子も持って いた。また、一部のサンプルでは、長時間日光に当たると肝斑を発症するリスクが高いことも分かっている。[25][28][2] 遺伝子データからは、縄文人は遺伝的に低身長になりやすく、また中性脂肪や血糖値が高く、肥満のリスクが高いことも示されている。一方で、飢餓に対する耐 性も備えていた。現代の日本人も、頻度は低くばらつきがあるものの、縄文時代の人口とこれらの対立遺伝子を共有している。これは、現代の日本人における混 血の推定値と一致している。Watanabeら(2023)は、日本人の低身長に対する遺伝的素因は、しばしば高い縄文人の祖先と相関し、その逆は高い大 陸東アジアの祖先と相関すると述べている。 劉ら(2024)は、現代の日本人における縄文人の混血率は地域によって異なり、混血率が最も高いのは日本の南部、特に沖縄(28.5%)で、次いで東北 (19%)と 西日本(12%)の順であると述べている。[29] 過去の研究では、東北、関東、九州の住民は琉球の住民と遺伝的により関連性が高く、近畿と四国の住民は大陸の東アジアの住民とより関連性が高いことが示さ れている。[25] ハプログループ ハプログループD-M55(D1a2a)とC1a1は、日本の縄文時代の歴史的人物の間で頻繁に見られたと考えられている。O-M119も、少なくとも一 部の縄文時代の遺跡で存在していたことが示唆されている。礼文島から発掘された3,800年前の縄文人の男性はハプログループD1a2b1(D-CTS 220)に属することが分かっている。[30] 現在、ハプログループD-M55は現代日本人の約35%[31]、ハプログループC1a1は約6%に見られる。D-M55は日本人(アイヌ人、琉球人、大 和人)にのみ、また、はるかに低い頻度ではあるが、韓国人にも見られる。D-M55はミクロネシアの5.1%、ティモールの0.2%、 中国 0~0.4%でも見られるが、これは最近になって混血したためであり、日本帝国(1868年~1945年)がそれらの地域を占領していた時代にまで遡るも のである。ハプログループC1a1は、現代の日本人の約6%に定期的に見られる。それ以外では、韓国、北朝鮮(黄海南道)、中国(寧安の朝鮮族、および凌 海、管城回族区、海港区、および定海区の漢族)の個人において散発的に観察されている。[33] 2021年の研究では、D-M55クレードの頻度は縄文時代後期に増加したと推定されている 。D1a2-M55とD1a-F6251亜系統(後者はチベット人、その他のチベット・ビルマ系民族、アルタイ人に多く、その他にも東アジア、東南アジ ア、中央アジアに中程度分布している)の分岐は、チベット高原付近で起こった可能性がある。 縄文人のミトコンドリアDNAハプログループの多様性は、ハプログループM7aとN9bの存在によって特徴づけられる。2004年と2007年に発表され た研究では、現代の日本人におけるM7aとN9bの合計頻度は、主流派の日本人では12~15%から17%であることが示されている。[35][36] N9bは北海道の縄文人に多く見られる一方、M7aは本州の縄文人に多く見られる。[37] しかし、 N9bは本州の縄文人の間で非常に低い割合でしか発見されていない。[37] M7aは、主に日本(縄文人を含む)、東アジアの他の地域、東南アジアで発見されているクレードのM7bと最も最近共通の祖先を共有していると推定されて おり、現在から33,500年(95% CI 26,300 <-> 38] ハプログループM7aの現存するメンバーはすべて、最も近い共通祖先を今から20,500年(95% CI 14,700~27,800年)前に共有していると推定されている。[38] ハプログループM7aは現在、沖縄で最も高い頻度で存在している。 形態学的特徴 Skull_of_Jomon_people_man.jpg 縄文時代晩期の男性頭蓋骨(レプリカ)。岩手県宮野貝塚出土。国立科学博物館にて展示。[39] 日本列島の各地から発掘された多数の縄文人の骨格標本に関する複数の研究により、日本の縄文時代の人口についてより詳しく知ることができるようになった。 縄文人は、他の東アジアの人々と比較的近かったが、アメリカ先住民の標本との類似点の方が多かった。 日本国内では、異なる縄文人の遺骨の間で地域差が認められた。 歴史的には、縄文人はモンゴロイドに分類されていた。[40][41] Jomon_people_Skull_and_Restoration_model_-_Niigata_Prefectural_Museum_of_History.jpg 新潟県立歴史博物館に展示されている縄文人の頭蓋骨の法医学的復元。 歯の形態学から、縄文人はスンダドント歯構造を有していたことが示唆されている。スンダドント歯構造は、現代の東南アジア人と台湾原住民に多く見られ、現 代の北東アジア人に多く見られるシノドント歯構造の祖先である。このことは、縄文人が現代の北東アジア人が形成される前に、一般的な「東アジア人祖先」か ら分岐したことを示唆している。 近藤ら(2017年)は、縄文時代の日本人の地域的な形態学的および頭蓋計測学的特徴を分析し、形態学的に多様であり、東北から西南西にかけての地域差が 認められることを発見した。 違いは頭蓋指数に基づいており、北海道の縄文後期の頭蓋は中頭蓋で、沖縄の頭蓋は短頭蓋であった。彼らは、「縄文人の頭蓋骨、特に神経頭蓋骨は、日本列島 全体にわたって東北から西南西への地理的傾斜が明瞭に認められ、両極端に位置する北海道と沖縄のサンプルが存在する」と結論付けた。 慎重を期して、以下の仮説を立てることができる。(a) 縄文人の形成は、西日本(沖縄や九州地方)ではなく、東日本または中部日本において進んだと思われる。(b) 九州の縄文人は、小規模で孤立した人口の歴史を持つ可能性がある。(c) 北海道の縄文人の人口の歴史は、深く根付いており、かつ/または、長期的な外部からの遺伝子流入の影響を受けている可能性がある。また、頭蓋長における地 域差は、遺伝的、系統発生的、および環境的影響に基づくものであると示唆している。[43] しかし、Gakuhariら(2020年)は、北海道と本州の縄文人は他のグループを排除してクラスターを形成し、まとまっていることを明らかにしてい る。[2] チャターズらによると、縄文人はアメリカ先住民のケネウィック・マンと類似点があるという。[44] チャターズは人類学者C. ローリング・ブレイスの言葉を引用し、縄文人とポリネシア人をひとつの頭蓋顔面「縄文 - 太平洋」クラスターに分類した。[45] チャターズはパウエルを引き合いに出し、縄文人はアメリカ先住民のケネウィック・マンとポリネシア人に最も似ていると主張している。彼によると、アイヌ人 は縄文人から派生した東アジアの民族であり、「西ユーラシアの人々よりも東南アジアの人々との生物学的類似性が高い」という。[44][46] パウェルはさらに、歯の分析により縄文人がスンダドントタイプであることが示されたと詳しく説明している。[45] 松村ら(2019年)によると、縄文人は 「長頭、大きな頬骨、著しく突出した眉間と眉弓、鼻根のくぼみ、低く幅広い顔」など、先史時代の中国南部や東南アジアの人々と類似した表現型を共有してい た。[47] 王ら(2021年)によると、これらの人々は表現型の不連続性にもかかわらず、後の東アジア人とも関連していた。[48] 縄文人の頭蓋顔面の特徴は、アイヌ人と沖縄人/琉球人にかなり受け継がれていた。[49] 前者は「ヨーロッパ人の顔の構造に関連する」2つの遺伝子を持っているが、東アジア人にみられる毛髪や歯の形態も有していた。[50] 顔の平坦さに関しては、 平坦さに関しては、アイヌ人はコーカソイドとモンゴロイドの中間であった[51]が、別の研究ではモンゴロイドの範囲内であるとしている[52]。後者に は「はっきりとした、平坦でない上顔面」があり、これは眉間と鼻根が突出していることで特徴づけられる[49]。 ATLレトロウイルス 縄文人によく見られる遺伝子は、ATL(成人T細胞白血病)の原因となるレトロウイルス(ヒトTリンホトロピックウイルス、HTVL-I)である。このウ イルスは成人T細胞白血病(ATL)の原因として発見され、京都大学ウイルス研究所の日沼拓夫氏によって研究が進められた。 日本にはウイルス保有者が多いことが知られていたが、東アジアの近隣諸国ではまったく発見されていない。一方、アフリカ人、アメリカ原住民、チベット人、 シベリア人、ビルマ人、ニューギニア原住民、ポリネシア人などには多く発見されている。日本での分布を見ると、特に九州南部、長崎県、沖縄、アイヌに多く 見られる。また、四国南部、紀伊半島南部、東北地方太平洋側(三陸)、隠岐諸島でも中程度の頻度で認められる。 全体として、ATLレトロウイルスの保有者は、辺地や離島に多いことが分かった。九州、四国、東北の各地域でATLが発症している地域を詳しく調べたとこ ろ、周囲から孤立し交通の便が悪い小規模な集落でキャリアの割合が高いことが分かった。 このウイルスの自然感染経路は、女性と子供の間での垂直感染(ほとんどが母乳を介したもの)と、男性と女性の間での水平感染(ほとんどが性交を介した男性から女性への感染)に限られている。[53] 以上のことから、樋沼は、このウイルスの高頻度領域は縄文人の高密度の遺跡を示すと結論づけた。[54] 他の集団への貢献 歴史的な集団 サハリンのオホーツク文化の遺跡の全ゲノム解析により、その遺跡は主に3つの集団、すなわち古代北東アジア人、古代旧シベリア人、および日本の縄文人から 派生していることが判明した。混血分析により、それらの人々はそれぞれ、約54%の古代北東アジア人、約22%の古代旧シベリア人、および約24%の縄文 人の祖先を持つことが明らかになった。 朝鮮半島南部の古代の遺跡の遺伝子解析では、縄文人の血統が約37%であることが明らかになった。一方、日本にある弥生人の遺跡では、縄文人の血統 (35~60%)と古代北東アジア人の血統(40~65%)がほぼ同量であることが判明した。これらの結果は、朝鮮半島に縄文人と同様の集団が存在し、初 期の日本語話者の形成に大きく貢献したことを示唆している。このように、先史時代の日本の「農業への移行は、土着の縄文人とムムン/弥生時代の本土アジア からの移住者とのほぼ同等の遺伝的貢献による同化のプロセスであり、置き換えではなかった」のである。[4][56] 現代の集団 アイヌ民族は、縄文時代由来の祖先型を最も多く示す現代の集団のひとつである(山本多助氏 36歳ごろ)。 縄文時代由来の血統は、日本列島全体で一般的に見られ、現代の日本人では約15%、琉球人では約30%、現代のアイヌ民族では約75%に及ぶ。また、ニブ フ族やウルチ族などの周辺集団では頻度が低く、 、また、朝鮮人やその他の沿岸部族にも見られ、縄文人が完全に他の集団から孤立していたわけではないことを示唆している。[2][28][55][57] [21] 王と王(2022年)は、三国時代に縄文人の血筋を持つ人々は、現代の韓国人の遺伝的構成に大きく貢献したと指摘している。しかし、この血統は、その後の 北方漢民族の到来により、時代とともに「希釈」された。[56] マッコールら(2018年)は、現在の東南アジア人は少なくとも4つの古代の集団に由来する祖先を持つと述べている。そのうちの1つは、古代日本人である イカワズ縄文人と祖先を共有する、ベトナム本土のホアビン人である。また、現在のアンダマン人のオンゲ族やマレー人のジェハイ族とも祖先を共有している。 [58] しかし、ヤンら(2020年)によると、縄文人の個体はシベリアと東南アジア南部のいくつかの沿岸の新石器時代の人々との類似性を示しており、後者は現在 の東南アジア南部の人々とグループ化されている。[59] 現代の日本人、ウルチ族、ニブフ族とともに、現在の台湾原住民(アミ族やタイヤル族など)も、日本本土の2500年前の人物と過剰な対立遺伝子共有を示しており、その人物は「典型的な縄文文化」の特徴を持っていた。[2] |
| In popular culture Aspects of the Jōmon culture and pottery were used in the video game The Legend of Zelda: Breath of the Wild. Nintendo's art director Takizawa Satoru said that the Jōmon culture was the inspiration for the "Sheikah slates, shrines and other ancient objects" in the game.[60] A recreated Jōmon village in the form of an experience park (Sarashina no Sato), which offers different activities, can be visited in Chikuma, Nagano.[61] |
大衆文化において 縄文文化や縄文土器の一部が、ビデオゲーム『ゼルダの伝説 ブレス オブ ザ ワイルド』で使用された。任天堂のアートディレクターである瀧沢悟は、ゲーム内の「シーカー石板、祠、その他の古代の遺物」は縄文文化から着想を得たものであると語っている。 さまざまなアクティビティが体験できる縄文時代の村を再現した体験型公園(更科の里)が長野県千曲市にあり、訪れることができる。[61] |
| History of Japan Yayoi people Okhotsk culture Satsumon culture Emishi Indigenous peoples of the Americas |
日本の歴史 弥生人 オホーツク文化 サツモン文化 エミシ アメリカの先住民 |
| https://en.wikipedia.org/wiki/J%C5%8Dmon_people |
★以下は日本語(ウィキペディア)のエントリー:「縄文人(じょうもんじん)」
| 縄文人(じょうもんじん)とは、縄文時代(新石器時代)に日本列島全域に居住していた人
々の総称。約1万6000年前から約3000年前まで現在の北海道から沖縄本島にかけて住み[1]、縄文文化と呼ばれる文化形式を保持していた。 |
・縄文人は、時間的(時代的)区分における「日本の先住性」をもった人
々のと解説される ・いつ:約1万6000年前から約3000年前(1万3000年のあいだ) ・どこ:現在の北海道から沖縄本島(敗戦後の日本のテリトリーに合致) |
| 概略 旧石器時代後の、約1万6,500年前の紀元前145世紀から約3,000年前の紀元前10世紀にわたる縄文時代の文化は、概ね現在の日本に分布してい た。そのため、この地域に居住していた縄文土器を作る新石器時代人を縄文人と見ることが出来る[2]。平均身長は男性が160cm弱、女性は150cm弱 でがっしりとしており、彫りの深い顔立ちが特徴で、世界最古級の土器を作り、約5000年前の縄文中期には華麗な装飾をもつ火焔(かえん)土器を創り出す など独自の文化を築いた[1]。東南アジアに起源をもつ人々ではないかと考えられてきたが、父系・母系両方の遺伝情報が入った核DNA解析の結果(三貫地 貝塚人骨、船泊遺跡縄文人骨)、東ユーラシアの人々の中では遺伝的に大きく異なる集団であることが判明した[1]。北海道や東北の縄文人は、本州や九州の 縄文人とは明らかに異なっており、本州と九州の縄文人は、現代の東アジア人に似ていたとされる[3][4]。 なお、もともと新石器時代という概念はヨーロッパを対象とした考古学における概念で農耕の存在を重視するものだったため、1960年代からしばらくの間は 縄文文化は新石器文化に分類されていなかった。 この縄文人は時期によって異なるが地域ごとに4から9のいくつかの諸集 団に別れていたと考えられている[5]。日本列島(旧石器時代のこの海域は後述のよ うに、現在とは相当に異なった海岸線を持っていた)に居住していた後期旧石器時代の人々が、後に縄文文化と総称される文化形式を生み出し、日本における縄 文人諸集団が出現したと推測されている。 |
・第一パラグラフでは、いわゆる「人種的」特徴が述べられる。 ・第二パラグラフでは、新旧の石器時代を、農耕の文化から峻別する。 ・「縄文時代」では「旧石器時代(非定住狩猟採集社会)と縄文時代の違いとしては、 土器と弓矢の使用[1]、磨製石器の発達[1]、定住化の始まりと竪穴建物・掘立柱建物の普及、環状集落等の定住集落や貝塚の形成、植物栽培(半栽培)の 始まりなどが挙げられる。また、遅くとも後期には稲作が開始されていたと考えられるが、多様な生業の一つに留まっていた点において、稲作に特化した弥生時 代とは異なるとされる」と説明されている。 ・第三パラグラフで興味ふかい記述は最初の文であり、縄文人はひとつの人間集団ではなく、複数のグルーに分かれていたことが示される。つまり、なぜ、この 項目の作成者は「縄文人」を、これらの複数の上に、ひとつのカテゴリー(=当時から現在も、存在も実在もしない「日本人」=現在の日本の政治的空間に居住 する人たち)で、むりやり、プロクステスの寝台のように、まとめた結果なのか? |
形質的特徴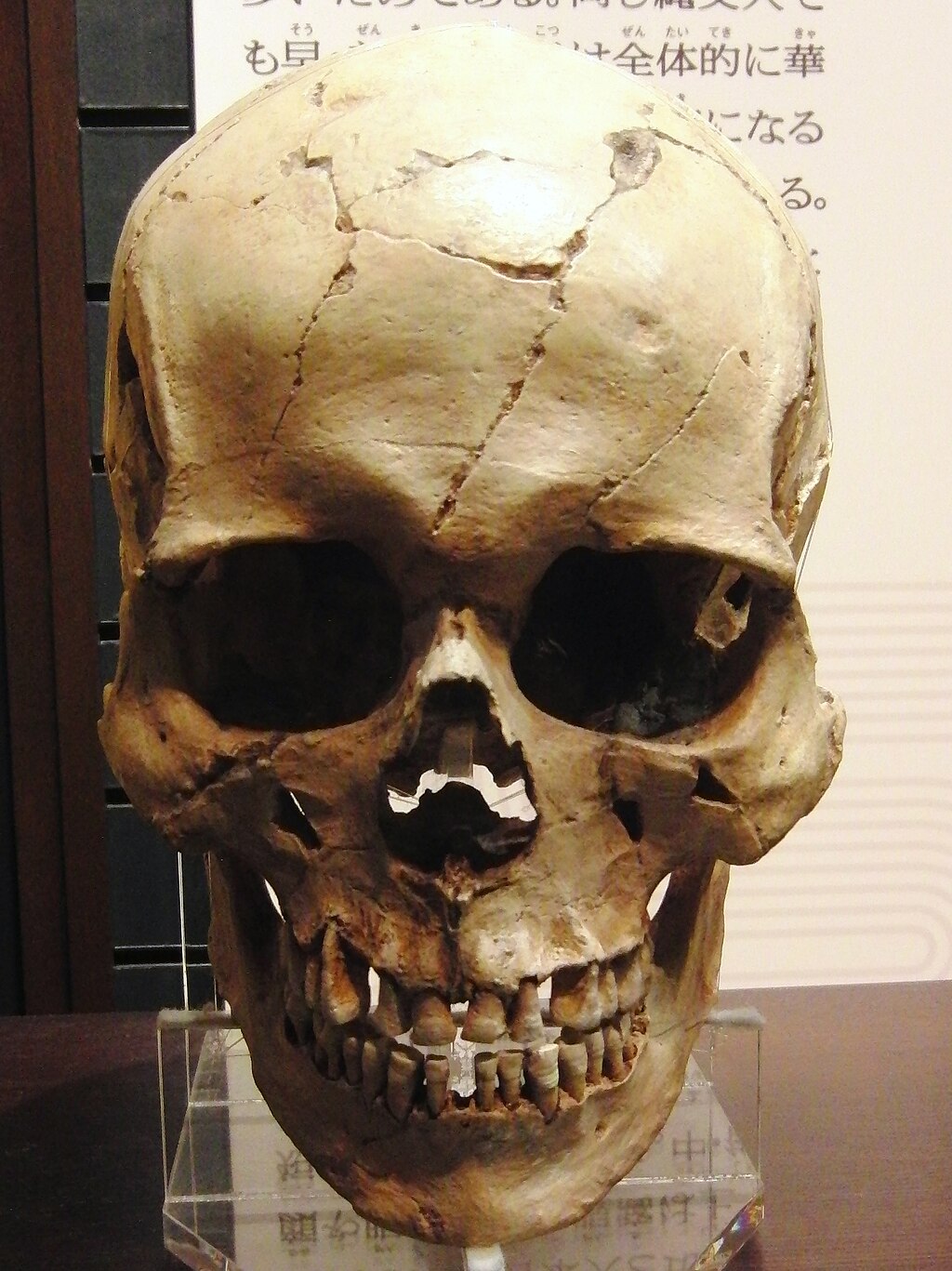 縄文時代晩期人の男性の頭骨(レプリカ)。宮野貝塚(岩手県)出土。国立科学博物館の展示[6]。 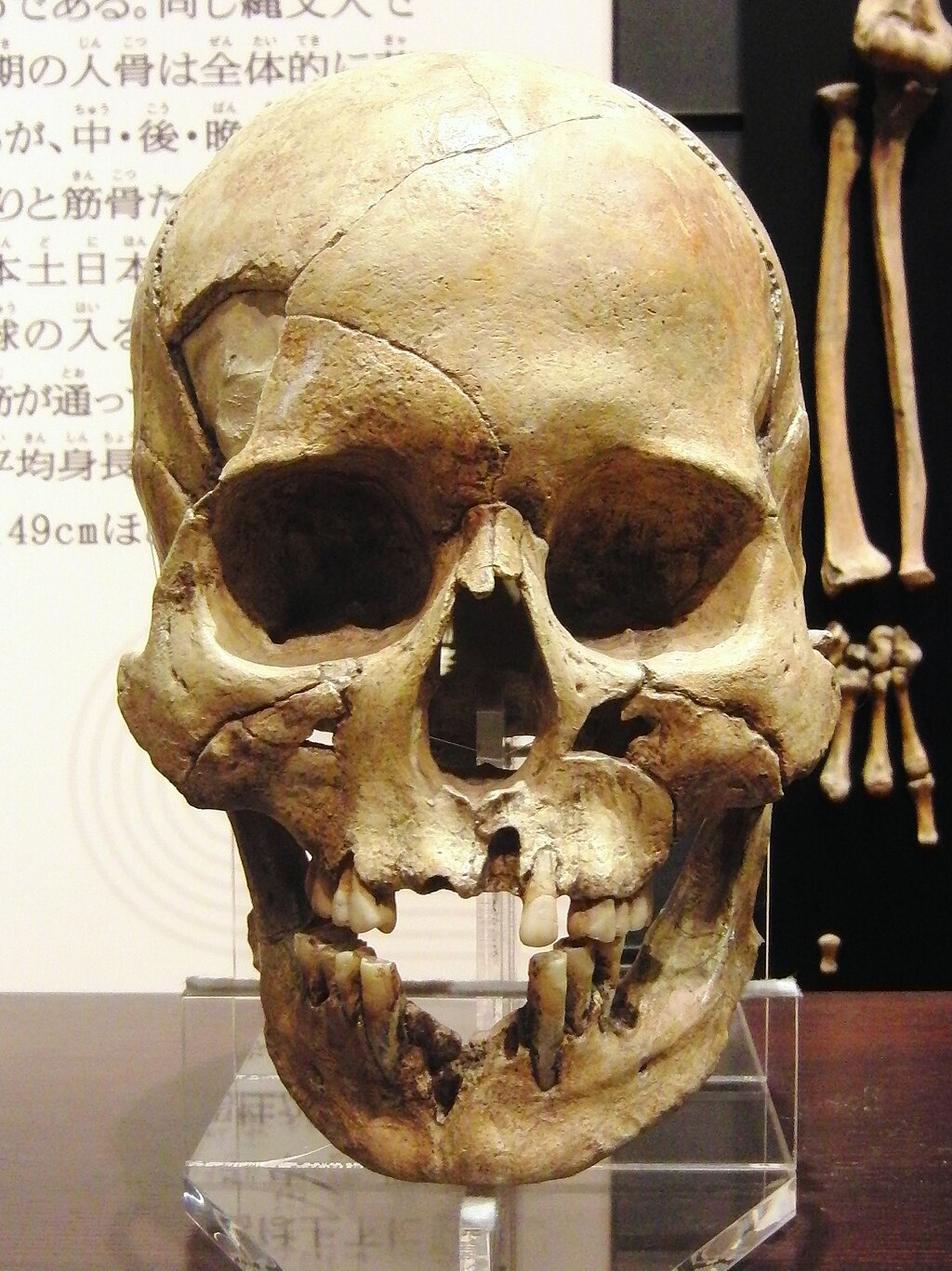 縄文時代後・晩期人の女性の頭骨(レプリカ)。蝦島貝塚(岩手県)出土。国立科学博物館の展示[6]。 彫りが深くて二重まぶた、眉や髭が濃い、沖縄県民やアイヌのよう な「濃い顔」が特徴。耳垢が湿っている。下顎が頑丈。がっしりしていて、平均身長は男性で160cm弱[7]。 こうした特徴を持つ人々が日本列島に出現した時期は、最終氷期の最寒冷 期(紀元前160世紀すなわち1万8000年前に氷河が堆積して海水面が最も低くな り、オホーツク海から北海道に歩いて渡れるようになった時期)が終わった後と見られる。ただし、既に日本列島に居住していた後期旧石器時代人の形質が変化 したものなのか、列島外から移住してきた人々の影響があるのかは不明である[8]。 3,800年前の縄文人女性の全ゲノム解析によると、縄文人サンプル は、太陽の下で長時間過ごすと肝斑を発症するリスクが高く[9][10]、同じく 2018年に北海道の礼文島から出土した3800年前の縄文人女性のゲノム情報をもとに顔を復元したところ、その女性の肌の色は現代の日本人よりもやや黒 く、髪は細く繊細で、目の色は明るい茶色だった[11]。 2015年に科学雑誌ネイチャー に掲載されたジナムらによるゲノムワイド一塩基多型(SNP)データ比較を用いた研究では、北海道の縄文人サンプルは、ヨーロッパ人(そしておそらく他の 集団においても)の顔の特徴に影響を与えることが以前の研究で知られている2つの遺伝子を含むいくつかの遺伝子座で本土の日本人とは異なることが判明した [12]。 近藤ら(2017)は、日本の縄文時代集団の地域的な形態学的・ 頭蓋計測学的特徴を分析し、それらが形態学的に異質であり、東北から南西に沿って連続的に 差異を示すことを明らかにした。彼らは、「縄文時代の頭骨は、特に脳頭蓋において、日本列島を横断する北東から南西への地理的な連続変異を 識別可能なレベ ルで示し、北海道と沖縄のサンプルはその両極端に位置する、とし、以下のシナリオは、注意深く仮定することができると結論づけた:(a)縄文人の集団形成 は、西日本(沖縄や九州地方)ではなく、東日本または中部日本で進行したようである。(b)九州の縄文人は、小規模で孤立した集団の歴史を持っている可能 性がある。(c)北海道の縄文人の集団の歴史は、深く根を下ろし、か つ/または長期的な外来遺伝子の流れの影響を受けた可能性がある[13]。 |
【人権上の配慮】 レプリカを含めて遺骨の展示は、先住民とその末裔の人たちに対して敬意を払うべきです。なぜなら、特定の民族集団の形態的特徴が、博物館で展示に値すると 考えられ、展示される人たちについての人権が配慮されることがなかったからです。この画面をクリックするとオリジナルの画面を見ることがことできます。 The display of the remains, including replicas, should be respectful to the indigenous people and their descendants. The human rights of the people whose remains are exhibited have never been taken into consideration, because the morphological characteristics of certain ethnic groups were considered worthy of museum display. Click on this image to see the original. ・「沖縄県民やアイヌのような「濃い顔」が特徴」→琉球人や沖縄人ではなく「沖縄県民」という地方行政区分と「アイヌ」のような民族ないしは「人種(的概 念)」が並存している。 ・縄文人も「渡来」の可能性→「 こうした特徴を持つ人々が日本列島に出現した時期は、最終氷期の最寒冷期(紀元前160世紀すなわち1万8000年前に氷河が堆積して海水面が最も低くな り、オホーツク海から北海道に歩いて渡れるようになった時期)が終わった後」 ・他方で、固有の後期旧石器時代人が縄文人に変化した可能性も示唆するが、最終的にわからないと言う→「既に日本列島に居住していた後期旧石器時代人の形 質が変化 したものなのか、列島外から移住してきた人々の影響があるのかは不明」 ・つまり、縄文人の起源は、2つあり、1)現在の日本列島にいた旧石器人の末裔が変化をとげたもの、2)日本が大陸と陸続きであったときに「渡来」したも のである。 近藤ら(2017)の研究では、縄文人は3つの系統にわかれる (a)縄文人の集団形成 は、西日本(沖縄や九州地方)ではなく、東日本または中部日本で進行したようである。 (b)九州の縄文人は、小規模で孤立した集団の歴史を持っている可能 性がある。 (c)北海道の縄文人の集団の歴史は、深く根を下ろし、かつ/または長 期的な外来遺伝子の流れの影響を受けた可能性がある[13]。 13. OSAMU KONDO, HITOSHI FUKASE, TAKASHI FUKUMOTO (2017). “Regional variations in the Jomon population revisited on craniofacial morphology”. Anthropological Science (日本人類学会) 125 (2): 85-100. doi:10.1537/ase.170428. ISSN 0918-7960. NAID 130006086850. 次のコラムで以下のような記述がある 瀬口(2014)によると、先史時代の縄文人は、日本へのさまざまな移 動ルートを使用して、多様な旧石器時代の人口から派生した[4]。 4. SCHmidT, Ryan W and Seguchi, N (2014). “Jomon Culture and the peopling of the Japanese archipelago: Advancements in the fields of morphometrics and ancient DNA”. Japanese Journal of Archaeology 2 (1): 34-59. |
| 縄文人のルーツ 形質人類学から見た縄文人のルーツ かつてアイヌはコーカソイドではないかと考えられて いたことがあるように、縄文人も形質的にコーカソイドに類似するとの研究[14] もあるものの、形質人類学の分野では、化石人骨が比較的多く見つかっている北東アジアと日本列島の旧石器時代人との類似を指摘する研究が多い。 これまでに出土した化石人骨と縄文人の関係を見ると、縄文人に最も近いとされているのは沖縄島出土の港川人(およそ1万8000年前)であるが、形質面か ら見ると縄文人は港川人の次の段階とまでは言えず、両者の間には更に1つか2つのミッシングリンクがあると考えられている[15]。また、港川人の頭骨は ワジャク人に近く、柳江人や山頂洞人(中国)にはそれほど似ていない為、少なくとも琉球弧の縄文人の祖先は環太平洋方面から来たのではないかとの見方があ る[15]。 一方で、縄文人は旧石器時代のシベリア南部の集団に由来するとする研究もある[16][4]。 瀬口(2014)によると、先史時代の縄文人は、日本へのさまざまな移 動ルートを使用して、多様な旧石器時代の人口から派生した[4]。 4. SCHmidT, Ryan W and Seguchi, N (2014). “Jomon Culture and the peopling of the Japanese archipelago: Advancements in the fields of morphometrics and ancient DNA”. Japanese Journal of Archaeology 2 (1): 34-59. この点で、縄文の生物学的アイデンティティは異質であり、縄文として知られている共通の文化に属している可能性のある多様な人々を示している可能性があり える。—Seguchi 2014 2017年の近藤修による頭骨測定の研究では、縄文人はかなり異質であることがわかった。「南縄文人」と「北縄文人」は大きく異なっていた。九州、四国、 本州の「縄文人南部」は、現代の東アジア人(「モンゴロイド」)とほとんど同じだったが、「縄文人北部」は異なる表現型を持っていた。縄文人の大多数は古 代東アジア人の子孫だが、北海道の縄文人は異なる遺伝的祖先を持っているようである。北海道縄文人と本州縄文人の間には遺伝子流動の証拠がある[13] 分子人類学から見た縄文人のルーツ 父系のルーツ 父系のルーツを辿ることができるY染色体ハプログループは、数万年にわたる長期的な追跡に適しており、1990年代後半から研究が急速に進展した。日本人 はY染色体ハプログループD1a2aの縄文系とハプログループO1b2の弥生系が最多の頻度を示し、埴原和郎らが唱えた「二重構造モデル」とも一致する結 果となった。Y染色体ハプログループD1a2aは現代の日本の民族集団(大和民族、琉球民族とアイヌ民族)に見られ、日本列島の本土と奄美群島・琉球列島 及び千島列島の3地域に多く見られるタイプである。また、Y染色体ではなくゲノムワイドの解析にもとづき「三重構造モデル」を提議する説もある[17]。 (→「ハプロタイプ研究」「二重構造モデル」) 国外では韓国、ミクロネシア、ティモール島などで低頻度にみられる。 このハプログループD1a2aはアイヌ人の75%に見られることから、D系統はかつての縄文人(旧石器時代のシベリア)のものであると考えられている。但 し縄文人のハプログループがD1a2aだけだった訳ではなくハプログループC1a1も縄文人由来と考えられている[18][19]。 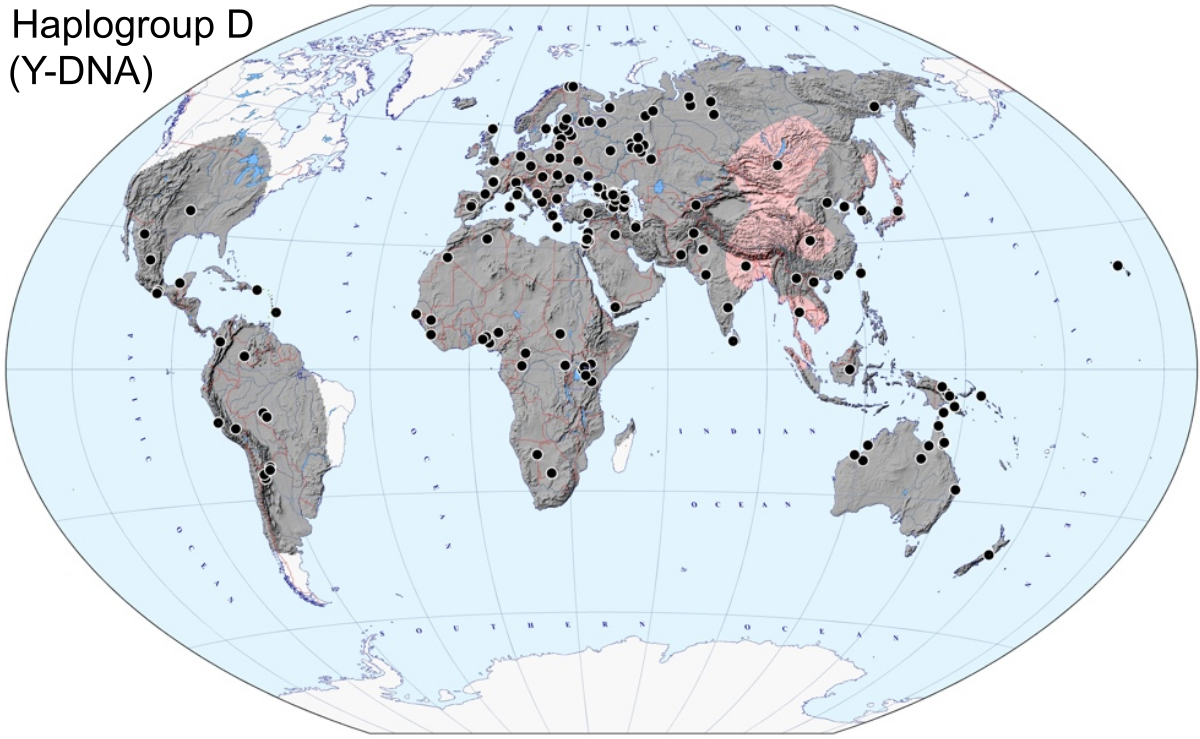 ハプログループDの分布 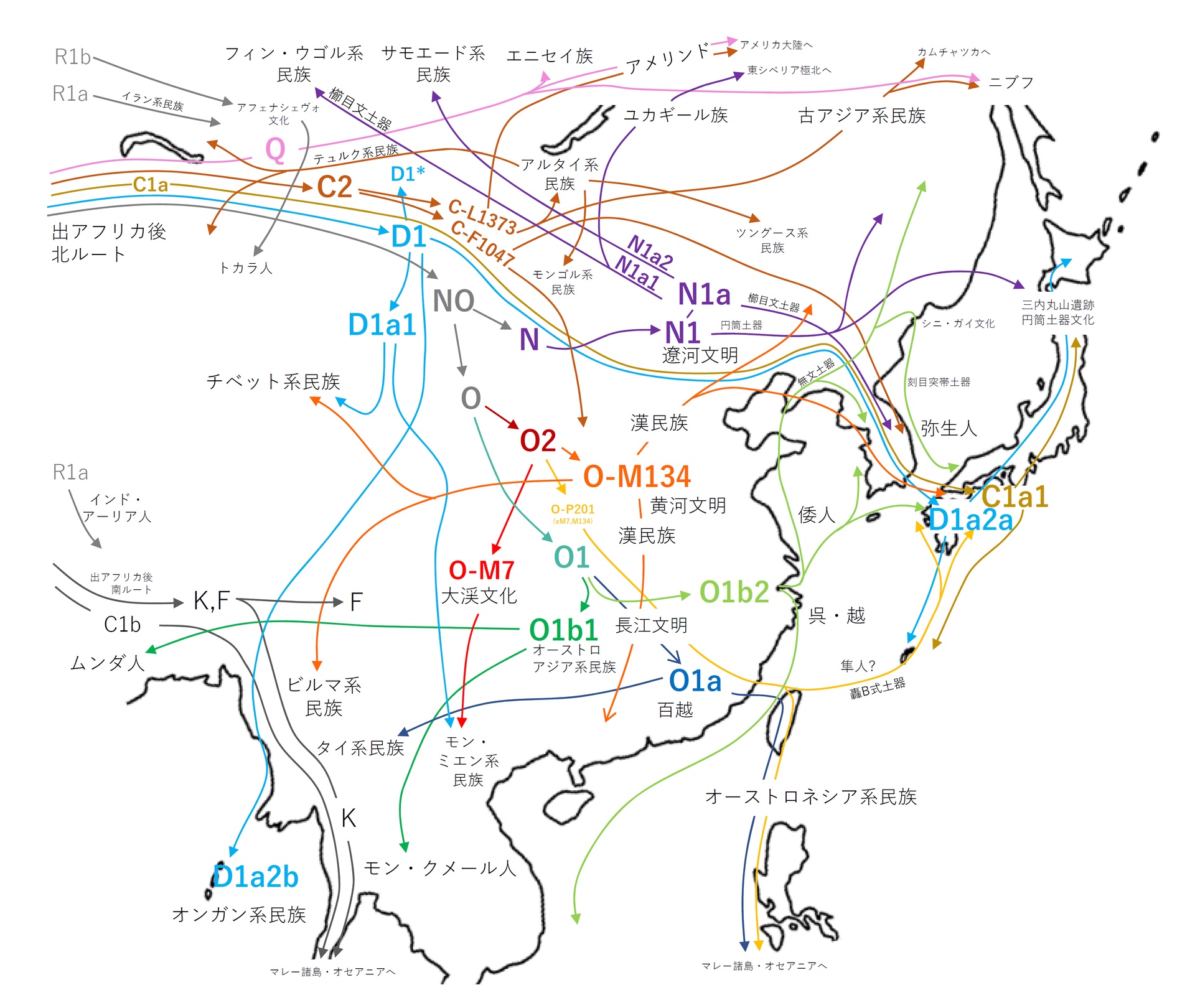 東アジアのY染色体ハプログループ移動図(→「ハプロタイプ研究」より) ハプログループD系統はYAP型(YAPハプロタイプ)ともいわれ、現代アジアにおいて支配的なO系統やC2系統とは分岐から7万年以上経ており、最も近 縁であり同じYAP型であるE系統とも6.5万年前に分岐した系統である。現在D系統は、日本列島以外で高頻度の地域はチベット[20][21] とアンダマン諸島[22] しかない。ハプログループDは、現代の東アジア人とシベリア人に近い「東アジアのハイランダーズ」(古代チベット人)に関連していて、縄文時代後期に増加 し、C1a1、K、Pなどの以前の縄文時代の系統に大きく取って代わったとされる[23][24]。 なお、当時の弥生人や現代東アジアにおいて支配的なO系統は、ウラル系のN系統やコーカソイド系において最多的なR系統などと近縁であり、縄文人やアイヌ 等のYAP型(D系統、E系統)とは全く異なるグループである。 以上のことから縄文人から自然進化的に弥生人が派生したという説は完全に否定されている。 日本列島にD系統の人々が入ってきたのは数万年前の最終氷期と考えられている。その証拠として、日本人のD系統にのみ見られる多くのSNPの発生があげら れる。SNPは突然変異により発生する確率的な事象であるから、発生数によって時間の経過が分かるのである。日本固有のD1a2a系統はその発生から 3.5-3.7万年ほどたっているとされ[25]、考古学から求められる日本列島に最初に現生人類集団が到来した時期と一致している。 長らく縄文人の人骨よりY染色体ハプログループは分析されてこなかったが、北海道礼文島の船泊遺跡(縄文時代後期前葉から中葉(約3,800~3,500 年前))から出土した人骨・船泊5号のY染色体ハプログループがD1a2a2a(D-CTS220)であることが判明した[26]。これにより「ハプログ ループD1a2aは縄文系である」という従来よりの仮説に一つ近づいたが、検証したのは長い縄文時代の中期のたった一人である為、時代、地域共に広範囲で の多数の検証が期待される。 とりわけハプログループC1a1は拡散年代と縄文文化開始の時期が一致しており、今後の研究いかんによっては初期の縄文人の主要なDNAとなる可能性があ る。C1a1は日本人固有であり、現在の日本ではおよそ5%の頻度で発見されている。 母系のルーツ 父系のルーツを辿れるY染色体ハプログループに対し、ミトコンドリアDNAハプログループは母系のルーツを辿ることができる。ただし、ミトコンドリア DNAは稀に男性のDNAが混じることや、人間より検証個体の多いネズミのDNA測定では、ハプログループの分岐や時期が事実とは全く異なっていたから、 あくまでもY染色体DNA等、他の資料と共に考察する必要がある。 ミトコンドリアDNA(母系)の分析によって縄文人のルーツの一角が解明され、日本固有のハプログループM7aや南方系と共通の遺伝子を持つハプログルー プBやFを持つことが知られている。宝来聡の研究によると、「東南アジアの少数民族から日本列島に位置する琉球弧人やアイヌまでが共通の因子を持つ」とさ れ、形質人類学においてはこれらの人々が縄文人と最も近いとされることから、縄文人のルーツは東南アジアの旧石器時代人との見方が可能である[27]。 これらを裏付けるように、国立科学博物館人類研究部 の篠田謙一らの研究では、鹿児島県霧島市・上野原遺跡の縄文人(25,000年前)から同様にハプログループM7aが検出され、縄文人は、現在は海底に沈 んでいる東南アジア・フィリピン沖のスンダランドが起源で、北上して南九州に到達し、大隅半島西北部の小高い台地にある上野原遺跡と呼ばれる「最古のム ラ」から日本列島全域へM7a系統の縄文人が拡散したと想定している。このM7a系統は、縄文時代にすでに北海道へも到達していたことが明らかとなってい る。 さらに2010年までに沖縄県石垣島の白保竿根田原洞穴遺跡(しらほさおねたばるどうけついせき)から発掘された、旧石器時代の人骨を国立科学博物館が分析した結果、国内最古の人骨(約2万- 1万年前)とされた4点のうち2点はハプログループM7aであることが明らかとなった[28]。 しかしながら、溝口優司は、5万年から6万年前にインドを経由し東南アジアで放散した東アジア人全体の祖先[29] の中から日本列島に到達したグループは複数存在し、東南アジアから北上する過程で台湾や南西諸島を経由し日本列島に到達した場合もあれば、一度北上し1万 5千年前にバイカル湖周辺で寒冷地適応した後に南下し朝鮮半島や中国から日本列島に移住した場合、バイカル湖を経由せずに大陸を海岸沿いに北上し、ブリ ヤートあたりから南下したルートが存在すると考えると、日本列島の遺伝的勾配をうまく説明できるという説を唱えている[30]。この説の要点は東アジアグ ループの成立年代が6万年前であり、バイカル湖で寒冷地適応したグループがアフリカから中央アジアを経由したわけではないので、宝来や篠田の説とも矛盾し ない。 また北海道の縄文人はハプログループN9bが最多でM7aは少なく、東北地方の縄文人も似た傾向を示している。このことから縄文人のルーツは一つではなく 複数あったと考えられる。 その他の遺伝子 縄文人の起源には複数の説がある。一部の説は東南アジアを、別の説は北東アジアを可能性のある起源であるとしている。一方、他の説は東アジアそのものを起 源としている。最新の遺伝学的研究は、縄文人はユーラシア大陸各地の集団によって形成された、としている[4][31][24]。2014年の遺伝学的及 び形態学的研究を概観した論文でも縄文人が複数の起源を持つ事が示唆されている[4]。 縄文人は、世界各地の集団と比較した場合、アフリカ人、ヨーロッパ人、サフール人(オーストラリア-メラネシア)、アメリカ先住民の集団よりは比較的東ア ジア人の集団に近い。ヒトゲノム多様性プロジェクト(英語: Human Genome Diversity Project)(HGDP)のデータを用いた全ゲノムに渡るSNP(一塩基多型)の比較においても、三貫地(さんかんじ)貝塚から出土した縄文人集団は現代東ユーラシ ア人全てと遠く離れており、その特異性が示された。東ユーラシア人内における三貫地縄文人集団の特異性は、ヨーロッパ人やアフリカ人と比べても同様であっ た。日本列島のアイヌ、本土日本人、琉球諸島の集団、そして北京の中国人(CHB28)を三貫寺の縄文人と比較した統計分析の場合、第一主成分でアイヌと 三貫寺縄文人が他の集団と分けられた。三貫地縄文人に最も近かったのはアイヌで、次いで琉球諸島の集団、そして本土日本人であった[32]。 今日ではほとんどの科学者は縄文人はユーラシア大陸の複数の集団の子孫であると考えている。標本の得られた縄文人の系統群は、東アジア人を含む他の人類集 団と遺伝的に距離がある。しかし、東アジアの沿岸部の集団との共通点も一部ある。その集団は本土日本人、ウリチ人、朝鮮人、台湾先住民である[33]。縄 文人は漢民族に代表される大陸のアジア人集団と比較的古い時期に分かれ、それは約3万8000年前から約1万8000年前までの間とみなすことができた。 縄文人は遺伝的には漢民族と比較した場合、日本列島のみならず、ロシア沿海州のウリチ族、朝鮮半島、台湾、フィリピンなど東アジアの沿岸部等の方に近いこ とが判明し、こうした要素を持つ人たちが大陸沿岸部に分散する中で一部が日本列島へ渡ってきた可能性が高そうだが、現状では日本列島の縄文人が大陸沿岸部 に広がった可能性も否定できない[26]。 26. HIDEAKI KANZAWA-KIRIYAMA and TIMOTHY A. JINAM and YOSUKE KAWAI and TAKEHIRO SATO and KAZUYOSHI HOSOMICHI and ATSUSHI TAJIMA and NOBORU ADACHI and HIROFUMI MATSUMURA and KIRILL KRYUKOV and NARUYA SAITOU and KEN-ICHI SHINODA (2019). “Late Jomon male and female genome sequences from the Funadomari site in Hokkaido, Japan”. Anthropological Science (日本人類学会) 127 (2): 83-108. doi:10.1537/ase.190415 縄文人(IK002)を含む古代および現在の個人の主成分分析(PCA)[34] 2020年に行われた全ゲノムの解析においては、縄文人の起源に関する更なる情報が示された。縄文人は様々なルートで日本に移住した様々な古代の人類集団 の子孫である事が分かった。縄文人は主に現在まだ標本の得られていない古代北ユーラシアの集団、そして様々な東アジアに関連した集団から形成されている。 この遺伝学的解析の結果が示すのは、旧石器時代に既に日本において異なる集団間の混血が起こり、その後も絶え間なく東アジア沿岸部からの遺伝子の流入があ り、その結果多様性のある集団となり、弥生人が到来する前に均質化した、という事である。チベット高原からの遺伝子の流入も検出され、ハプログループ D1a2a (Y染色体)と関連している。このグループは縄文時代後期になって初めて日本における優勢なハプログループとなった。KとFの系統は、縄文時代初期に存在 していたが、CとDに取って代わられたことが示唆されている。伊川津貝塚から出土した縄文人の標本一体、及びチベット高原の古代の標本の分析からは、両者 の間には部分的な共通祖先が発見されただけであった。これは古代チベット高原の集団及びチベット・ビルマ語派の集団に関連するハプログループDが拡散する 際に、正の遺伝的ボトルネックが起きたことを示している。遺伝的証拠から、ヒマラヤ山脈付近の東アジアの源流集団が、縄文時代の日本の集団に祖先をもたら したことが示唆され、古代の東南アジア人への祖先の寄与は少なかった。著者らは、このことは旧石器時代に中国南部または中部を通って日本へ内陸移動したこ とを示唆していると結論づけた [35][36]。 35.Yang, Melinda A.; Fan, Xuechun; Sun, Bo; Chen, Chungyu; Lang, Jianfeng; Ko, Ying-Chin; Tsang, Cheng-hwa; Chiu, Hunglin et al. (2020-07-17). “Ancient DNA indicates human population shifts and admixture in northern and southern China” (英語). Science 369 (6501): 282-288. doi:10.1126/science.aba0909. ISSN 0036-8075. PMID 32409524. 36.Boer, Elisabeth de; Yang, Melinda A.; Kawagoe, Aileen; Barnes, Gina L. (2020/ed). “Japan considered from the hypothesis of farmer/language spread” (英語). Evolutionary Human Sciences 2. doi:10.1017/ehs.2020.7. ISSN 2513-843X. 古代シベリアからの遺伝子流入は北海道の北方縄文人にも検出され、後に北海道から本州北部(東北)にも遺伝子流入した[35][36]。2021年、北海 道の古代の人口集団は、縄文人と旧石器時代の北ユーラシアに土着した「終末期後期旧石器人」(TUP人)から形成されたことが確認された。東アジアから紀 元前15,000年頃に渡来した縄文人と、それ以前に渡来した「終末期後期旧石器時代北ユーラシア人」とが融合して、北海道の縄文人が形成された [37]。2020年のガクハリらも同様に、古代北ユーラシア人(マリタ遺跡のMA-1サンプルによってサンプル化された)、または類似のグループから北 日本への遺伝子流入の可能性を指摘しており、これはシベリアの細石刃文化の導入と明らかに関連づけることができる、とした[34]。Cookeらによる 2021年の全ゲノム分析。現在分析されているすべての縄文人サンプルと他の集団との系統発生関係を再評価し、縄文人の系統は紀元前15,000年から2 万年の間に現代の東アジア人から分裂し、外部の集団から大部分が孤立したと結論付けた。著者らはその後、縄文人が後期旧石器時代の大陸の人々と接触したか どうかを分析した。分析によると、縄文人は、古代北ユーラシア人としても知られる、ヨーロッパに深く関係する北シベリアの旧石器時代後期旧石器時代のヤナ RHSサンプルに関連する集団から混合物を受け取り、最終氷期最盛期以前に北ユーラシアに広まった。現代の日本人の間で推定される縄文人の祖先は、平均 9.31%と推定されている[17]。 2020年にケンブリッジ大学出版局が発行したある研究では、縄文人は均質ではない集団であり、弥生人が移民するよりも古い紀元前6,000年前頃に北東 アジア人の移民があり、その集団が大平山本遺跡などの初期の土器文化に代表される縄文時代草創期の文化をもたらした、と示唆している(この文献は出典なし)。その研究者たちは更 に、弥生人の到来の前にオーストロネシア人が日本の南端(特に先島諸島)にいた可能性について言及している[38]。 38. Chaubey, Gyaneshwer; Driem, George van (2020/ed). “Munda languages are father tongues, but Japanese and Korean are not” (英語). Evolutionary Human Sciences 2. doi:10.1017/ehs.2020.14 2022年にメリンダA.ヤンが行ったアジアの人々に関する論文のレビューでは、縄文人は「東アジアおよび東南アジア」(ESEA)系統の他の人々と密接 に関係していることがわかった。このESEAの血統は、現代の東アジアおよび東南アジアの人々、シベリアの先住民、アメリカ大陸の先住民の祖先の一部であ り、そして古代のホアビニアン(英語版)の血統と田園洞人のサンプルを含んでいる。ただし、オーストラリア人やヨーロッパ人とは異なる。縄文人は、古代の 中国南部(Longlin)のサンプルと同様に、ホアビニアンの血統と田園洞人のサンプルと比べて、9,000-4,000年前の東アジアの北部と南部の 沿岸部からのサンプルに近いが、その中では外れた位置にある[39]。 39. Yang, Melinda A. (2022-01-06). “A genetic history of migration, diversification, and admixture in Asia” (英語). Human Population Genetics and Genomics 2 (1). doi:10.47248/hpgg2202010001. ISSN 2770-5005. HLA IおよびHLA II遺伝子とHLA-A、-B、および-DRB1遺伝子頻度の分析からは、アイヌとアメリカ大陸の先住民、特にトリンギットなどの太平洋岸北西部の集団と の類似性がみられ、アイヌ民族とアメリカ先住民のグループの主な祖先は、南シベリアの旧石器時代の集団にまで遡ることができると示唆された[40]。 縄文時代のさまざまなサンプルの人口ゲノムデータは、紀元前1万5000年頃に他の東アジアの人々から分岐したことを示している。しかし現在の典型的なモ ンゴロイドとは差異が大きい。また、縄文時代の様々なサンプルから得られた集団ゲノム学的データは、彼らが3万年から2万年前に他の東アジアの人々と分岐 したことを示している。紀元前15,000年から20,000年にかけて日本列島に移住した後、彼らは外部からの遺伝子の流入からほぼ隔離された[41] [42]。 41.Katsushi Tokunaga and Jun Ohashi and Makoto Bannai and Takeo Juji (2001). “Genetic link between Asians and native Americans: evidence from HLA genes and haplotypes”. Human Immunology 62 (9): 1001-1008. doi:10.1016/S0198-8859(01)00301-9. ISSN 0198-8859. 42.Cooke, Niall P.; Mattiangeli, Valeria; Cassidy, Lara M.; Okazaki, Kenji; Stokes, Caroline A.; Onbe, Shin; Hatakeyama, Satoshi; Machida, Kenichi et al. (September 2021). “Ancient genomics reveals tripartite origins of Japanese populations” (英語). Science Advances 7 (38): eabh2419. Bibcode: 2021SciA....7.2419C. doi:10.1126/sciadv.abh2419. PMC 8448447. PMID 34533991. またヨーロッパ人や古代北ユーラシア人に関連する北シベリアの遺跡(ヤナRHS遺跡(英語: Yana Rhinoceros Horn Site)から出土した人骨(Yana_UP)から検出された遺伝子が、分析された縄文人のサンプルで検出され、北海道の縄文人の北部のサンプルでより高 い頻度で検出された[17]。しかし、縄文人とヤナ人の間には同数値が検出されたが、わずかな有意差しかなかった。より質の高い古代サンプルを用いた更な る検証研究が必要である。今後の研究、特に古シベリア人と古代中央アジア人の追加サンプルは、ヤナ人と縄文人の間の潜在的な遺伝的つながりの理由を解明す るのに役立つと思われる[43]。 考古学から見た縄文人のルーツ 日本列島に居住した後期旧石器時代人を縄文人の直接の祖先と見た場合、問題となるのは、これら日本列島の後期旧石器時代人はどこから来たのかという点であ る。石器の形式に注目してみると、後期旧石器時代の日本列島にはナイフ型石器と細石刃という、同じ用途に用いられる2種類の石器が存在していた。 ナイフ型石器は大陸では出土例が無いもので、日本列島で独自に発達したものと考えられ、鹿児島県上野原遺跡の調査などから発見されている。 一方で、細石刃はバイカル湖周辺に起源を持つもので、日本列島に伝播したのはおよそ2万年前、宗谷海峡経由で北海道にもたらされた。細石刃が東北や北陸に 伝わるのはおよそ1万5000年前である。つまり、この時期に北東アジア方面から細石刃の技術を持った少数の人々(ハプログループC2(C-M217)ら が北方からも日本列島に移動してきたことになるという。また華北・朝鮮半島経由で九州に、さらには西南日本に伝播してきた細石器もあり、最古の出土物から 1.6-1.5万年前に伝播したものと考えられているが、もっと遡るという説もある[44]。 |
・現在の生物人類学研究では、肌の色の違いによる人種識別、例えばコーカソ
イドなどという表現は使わない(→「人種理論」) ・瀬口(2014)によると、先史時代の縄文人は、日本へのさまざまな 移動ルートを使用して、多様な旧石器時代の人口から派生した[4]。 4. SCHmidT, Ryan W and Seguchi, N (2014). “Jomon Culture and the peopling of the Japanese archipelago: Advancements in the fields of morphometrics and ancient DNA”. Japanese Journal of Archaeology 2 (1): 34-59. ・ この点で、縄文の生物学的アイデンティティは異質であり、縄文として知られている共通の文化に属している可能性のある多様な人々を示している可能性があり える。—Seguchi 2014 ・つまり、縄文人は、生物的遺伝的に多様でも、共通の文化的特質をもっていた。 ・北海道縄文人と本州縄文人の間には遺伝子流動 【遺伝子流動(gene flow、gene migration)】 ・「遺伝子流動(いでんしりゅうどう、gene flow、gene migration)とは、ある集団から別の集団への対立遺伝子、あるいは遺伝子の移動のこと。 集団の中、あるいは外への移住は、集団中の対立遺伝子頻度(ある遺伝的変異を持った構成個体の割合)を変化させる。場合によっては、ある種、集団からなる遺伝子プールに新しい遺伝的変異を加えることにもなるかもしれない[1]。 異なる集団間での遺伝子流動の比率に影響する要因は様々である。最も重要な要因の一つは生物の持つ移動性であり、一個体が高い移動性を持つほど、移住のポテンシャルは高くなる。平均的には、植物より動物が高い移動性を持つ傾向にある。 二つの集団間で維持される遺伝子流動は二つの遺伝子プールを結合し、集団間で比較した遺伝的変異を減少させる。遺伝子流動は、遺伝子プールを融合させることにより種分化とは正反対に働く力であり、種分化や姉妹種を生み出す原因である遺伝的分化を成立させないのだ。 例えば、道路の両脇に生えたイネ科植物を想像してほしい。植物の花粉は一方から反対側に飛び、逆もまた同様である。花粉は植物を受粉させ、子孫が成長する。その結果、子孫の花粉の対立遺伝子は反対側の集団から飛んできた対立遺伝子に置き換わっている。」 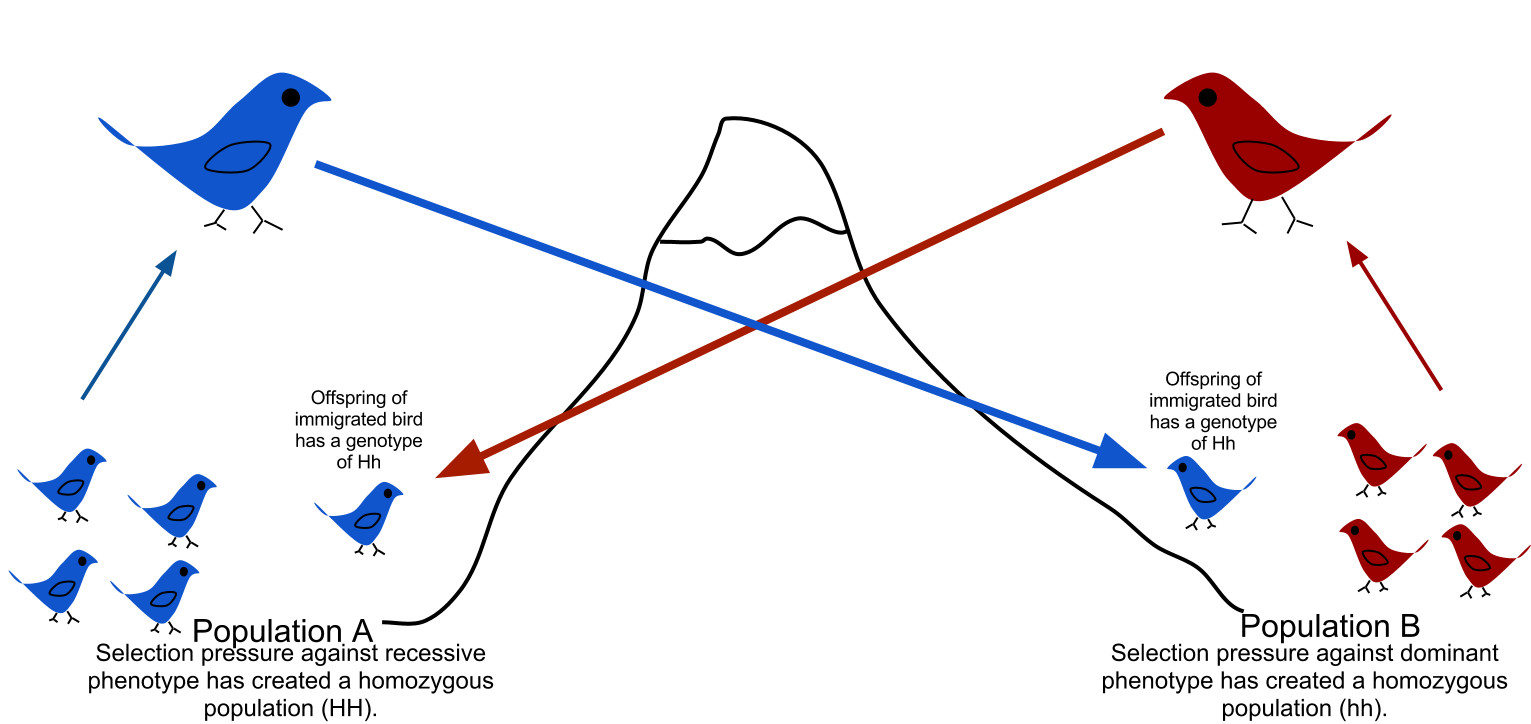 17. 三重構造モデル(tripartite origins of Japanese populations)? ・Cooke, Niall P.; Mattiangeli, Valeria; Cassidy, Lara M.; Okazaki, Kenji; Stokes, Caroline A.; Onbe, Shin; Hatakeyama, Satoshi; Machida, Kenichi et al..(2021) “Ancient genomics reveals tripartite origins of Japanese populations”. Science Advances 7 (38): eabh2419. doi:10.1126/sciadv.abh2419. ISSN 2375-2548. PMC 8448447 「【要約】先史時代の日本は、過去3000年の間に急速な変貌を遂げた。まず、採集から湿田稲作へ、そして国家形成へと。長年の仮説では、日本本土の集団 は縄文時代の狩猟採集民と弥生時代の農耕民の二重祖先から派生したと考えられている。しかし、農耕民族の移動とその後の社会文化的変化がゲノムに与えた影 響については、まだ不明な点が多い。我々は、農耕時代以前と以後の12種類の古代日本人ゲノムを報告する。その結果、縄文人は数千年にわたり1000人程度の小規模な有効人口を維持していたが、大陸の集団との深い乖離が見られたのは、海面上昇によって日本が島国化した2万~1万5000年前であることがわかった。稲作は北東アジアに祖先を持つ人々によって導入された。意外なことに、古墳時代には東アジアの祖先が流入していた。これら3つの祖先構成要素は、現在の集団を特徴づけており、日本人のゲノム起源に関する三者モデルを支持している。」 ・縄文人は、列島が陸続きの時代には、大陸との遺伝的かかわりをもっていたが、海面上昇以降は、独自の進化をとげた。また、縄文系はY染色体ハプログループD1a2aをもち、弥生系は「現代東アジアにおいて支配的なO系統」からなりたち縄文系の「YAP型(D系統、E系統)」とは異なる。「縄文人から自然進化的に弥生人が派生したという説は完全に否定」された。 ・26. HIDEAKI KANZAWA-KIRIYAMA and TIMOTHY A. JINAM and YOSUKE KAWAI and TAKEHIRO SATO and KAZUYOSHI HOSOMICHI and ATSUSHI TAJIMA and NOBORU ADACHI and HIROFUMI MATSUMURA and KIRILL KRYUKOV and NARUYA SAITOU and KEN-ICHI SHINODA (2019). “Late Jomon male and female genome sequences from the Funadomari site in Hokkaido, Japan”. Anthropological Science (日本人類学会) 127 (2): 83-108. doi:10.1537/ase.190415 「【要約】船泊縄文人は3500-3800年前に北海道の礼文島に住んでいた狩猟採集民である。本研究では、船泊縄文人の女性(F23)と男性(F5)か ら、それぞれ高深度核ゲノム配列と低深度核ゲノム配列を決定した。F23の核DNAを遺伝子型解析し、ヒト白血球抗原(HLA)クラスIの遺伝子型と表現 形質を決定した。さらに、F23とF5の両方でCPT1A遺伝子の病原性変異が同定された。この変異は高脂肪食の摂取に代謝上の利点をもたらし、その対立 遺伝子頻度は北極圏の集団では70%以上であるが、他の地域では見られない。この変異型は、陸上動物や海洋動物を漁猟していた船泊縄文人の生活様式に関連 している可能性がある。F23では高いホモ接合性(HBD)が観察されたが、10cM以上のHBDトラクトは非常に限られており、北方縄文人集団の集団規 模が小さいことが示唆された。我々の解析から、縄文人の集団規模は5万年前から減少し始めたことが示唆された。F23、現代/古代ユーラシア人、ネイティ ブ・アメリカンの系統関係は、おそらくネイティブ・アメリカンの祖先が東ユーラシア人から分裂する以前、40000年前の田園(洞人)が分裂した後に、東 ユーラシアでF23が深く分岐したことを示しており、北方縄文人は長い間、大陸の東ユーラシア人から遺伝的に隔離されていたことを示している。興味深いこ とに、現代の日本人、ウルチ族、朝鮮族、台湾原住民、フィリピン人の集団は、漢民族よりもF23に遺伝的に近いことがわかった。さらに、F5のY染色体 は、現代日本人の集団では稀なハプログループD1b2bに属していた。これらの発見は、東ユーラシアにおける古代人の集団構造の歴史と復元に関する洞察を 与えるものであり、F23ゲノムのデータは今後の研究のための縄文参照ゲノムとみなすことができる」。 ・35.Yang, Melinda A.; Fan, Xuechun; Sun, Bo; Chen, Chungyu; Lang, Jianfeng; Ko, Ying-Chin; Tsang, Cheng-hwa; Chiu, Hunglin et al. (2020-07-17). “Ancient DNA indicates human population shifts and admixture in northern and southern China” (英語). Science 369 (6501): 282-288. doi:10.1126/science.aba0909. ISSN 0036-8075. PMID 32409524. 「【要約】中国への、そして中国国内への人類の移動の歴史は、考古学的調査や現代人の遺伝学的研究だけから決定することは困難であった。Yangらは、 9500年から300年前の中国国内26人のDNA配列を決定した。これらの個体について、以前に塩基配列が決定された古代の個体や世界的な集団を代表す る現代のゲノムとともに解析した結果、中国北部と南部で古代の人類が分裂していることが示された。新石器時代の中国北部の個体は現代の東アジア人に最も近 いが、中国南部の古代の個体は現代の東南アジア人に最も近縁であり、現代のオーストロネシア系個体群との親和性を示している。これらの結果は、新石器時代 に民族の南下と混血があり、それが現代の東アジアの集団を生み出したことを示している。」 36.Boer, Elisabeth de; Yang, Melinda A.; Kawagoe, Aileen; Barnes, Gina L. (2020/ed). “Japan considered from the hypothesis of farmer/language spread” (英語). Evolutionary Human Sciences 2. doi:10.1017/ehs.2020.7. ISSN 2513-843X. 「【要約】形式的には、日本に適用される農耕・言語拡散仮説は、農耕の導入と日本語の普及(紀元前500年頃から紀元後800年頃まで)に関連している。 この仮説に関連して、遺伝学、考古学、言語学から得られた現在のデータを検証する。しかし、これらの学問分野の証拠基盤は異なる時代から得られている。遺 伝学的データは主に、稲作農耕が導入される前の、現在の日本人と先史時代の縄文人(紀元前14,000~300年)から得られている。考古学的には、西日 本の弥生時代(紀元前900年頃~紀元後250年頃)の農耕の証拠が最もよく知られているが、ここでの焦点である東北日本についてはほとんど知られていな い。また、先史時代の言語についてはかなりの仮説が立てられているが、歴史的な言語や方言の島々への広がりは、先史時代と関連付けるのは難しい。DNA分 析に利用可能な弥生人の骨格資料が不足しているため、農業以前の縄文人が稲作農耕民とどのように交流していたかを直接的に研究することは非常に困難である が、縄文人の遺伝学的考察は、彼らの関係をさらに研究するための舞台を整えるものである。現代の言語学的研究は、出雲(島根県)と日本海沿岸が、関東地方 を越えて農耕民が本州北東部に居住したことを考察する上で、思いがけない役割を果たしている。」 38. Chaubey, Gyaneshwer; Driem, George van (2020/ed). “Munda languages are father tongues, but Japanese and Korean are not” (英語). Evolutionary Human Sciences 2. doi:10.1017/ehs.2020.14 「【要約】20年以上前、ある集団で話されている言語の言語的親和性は、その集団で優勢な父系、すなわちY染色体系統と相関する傾向があることが観察され た。このような相関関係はどこにでも見られるが、普遍的なものではないことが判明した。言語系統地理学と遺伝系統地理学の相関関係のような顕著なパターン には顕著な例外があり、特に興味を引き、その解明が望まれる。オーストロアジア語族の中で、ムンダ諸語は明らかに父語のケースであるが、日本語と韓国語は 明らかに父語ではない。本研究では、ムンダ語と日本語のケースを並置する。まず、Y染色体O系統の系統地理を簡単に説明する必要がある。次に、三角測量に よって、長い間失われていた両者の民族言語史前史の輪郭と特殊性を明らかにする」。 ・縄文時代の様々なサンプルから得られた集団ゲノム学的データは、彼らが3万年から2万年前に他の東アジアの人々と分岐 したことを示している。紀元前15,000年から20,000年にかけて日本列島に移住した後、彼らは外部からの遺伝子の流入からほぼ隔離された. 42.Cooke, Niall P.; Mattiangeli, Valeria; Cassidy, Lara M.; Okazaki, Kenji; Stokes, Caroline A.; Onbe, Shin; Hatakeyama, Satoshi; Machida, Kenichi et al. (September 2021). “Ancient genomics reveals tripartite origins of Japanese populations” (英語). Science Advances 7 (38): eabh2419. Bibcode: 2021SciA....7.2419C. doi:10.1126/sciadv.abh2419. PMC 8448447. PMID 34533991.(→上掲17.と同じ論文) ・「【要約】先史時代の日本は、過去3000年の間に急速な変貌を遂げた。まず、採集から湿田稲作へ、そして国家形成へと。長年の仮説では、日本本土の集団は縄文時代の狩猟採集民と弥生時代の農耕民の二重祖先から派生したと考えられている。しかし、農耕民族の移動とその後の社会文化的変化がゲノムに与えた影響については、まだ不明な点が多い。我々は、農耕時代以前と以後の12種類の古代日本人ゲノムを報告する。その結果、縄 文人は数千年にわたり1000人程度の小規模な有効人口を維持していたが、大陸の集団との深い乖離が見られたのは、海面上昇によって日本が島国化した2万 ~1万5000年前であることがわかった。稲作は北東アジアに祖先を持つ人々によって導入された。意外なことに、古墳時代には東アジアの祖先が流入してい た。これら3つの祖先構成要素は、現在の集団を特徴づけており、日本人のゲノム起源に関する三者モデルを支持している」 |
| ATLのレトロウイルス 縄文人に関連する遺伝子として、ATLのレトロウイルス (HTLV-1) がある。このウイルスは成人T細胞白血病 (ATL) を引き起こす原因として発見されたもので、HTLVは京都大学ウイルス研究所教授の日沼頼夫によって研究が進められた。 日本人にはこのウイルスキャリアが多数存在することは知られていたが、東アジアの周辺諸国ではまったく見出されていない。いっぽうアメリカ先住民やアフリ カ、ニューギニア先住民などでキャリアが多いという特徴をもつ[45]。日本国内の分布に目を転じてみると、九州南部と長崎県に多いのが目立つ。そして沖 縄やアイヌに特に高頻度で見られ、四国南部、紀伊半島の南部、東北地方の太平洋側、隠岐、五島列島などの僻地や離島に多いことが判明した[46]。九州、 四国、東北の各地方におけるATLの好発地域を詳細に検討すると、周囲から隔絶され交通の不便だった小集落でキャリアは高率に温存されている。東京、大阪 など大都市で観察される患者の90%以上は九州などに分布するATL好発地帯からの移動者で占められていた。このウイルスの感染機構は生きた感染リンパ球 と非感染リンパ球の接触で起こり、空気や通常接触では感染せず、体液(血液、母乳、精液など)が主な感染源になる。自然感染の経路としては母児間の垂直感 染と男女間の水平感染に限られることになる。特に夫から妻への感染が多く逆はほとんど観察されない。 以上より、日沼はこのウイルスのキャリア好発地域は、縄文系の人々が高密度で残存していることを示していると結論付けた[47]。HTLVはかつて日本列 島のみならず東アジア大陸部にも広く分布していたが、激しい淘汰が繰り返されて大陸部では消滅したようである。弥生時代になってウイルス非キャリアの大陸 集団が日本列島中央部に多数移住してくると、列島中央部でウイルスが薄まっていったが、列島両端や僻地には縄文系のキャリア集団が色濃く残ったものと考え られる。 |
|
| 縄文人観の歴史的変遷 縄文時代の日本列島に住んだ人々に対するイメージは、その時々の日本社会の風潮と呼応して様々に移り変わってきた[48]。 「日本列島の先住民族」としての縄文人 明治期には縄文人は「石器時代人」と呼ばれ、日本列島の先住民族と考えられていた。この時期には日本人の祖先は「天孫族」と呼ばれており、記紀神話にある ように列島史のある段階で別の場所から日本列島にやって来た人々であるとされていた。その為、「石器時代人」はいわゆる日本民族の祖先ではなく、アイヌの 祖先あるいはアイヌ神話に登場するコロポックルではないかと考えられており、この論点を巡って「アイヌ・コロポックル論争」と呼ばれる論争も発生した。 また土偶がグロテスクに思われたため日本人の祖先としてふさわしくないとして否定されることもあった[49]。 先住民族とする見方は鳥居龍蔵による「固有日本人論」にも受け継がれたが、一方で昭和に入ると浜田耕作が縄文人を日本民族の祖先と見る説を発表し、学界に 一石を投じた。 「高級狩猟民」としての縄文人 1930年代には唯物史観が登場し、縄文人を経済面から新たに捉え直そうとする動きが始まる。代表的な論者として山内清男が挙げられる。山内は縄文人を、 男性が狩猟・漁労に従事し、女性が採集活動に従事するという分業体制を持った、発達した狩猟採集民族であったと考えた。 「弥生文化の母体」としての縄文人 戦後(第二次世界大戦後、太平洋戦争後)つまりまだDNA分析技術が無く、学者もまだ出土「史料」だけに基づいて起きた出来事を空想するにとどまっていた 時代には、それまで「弥生人に単純に置換された存在」と見られていた[要出典]縄文人を、「弥生文化を主体的に受容して弥生人へと変化していった人々[要 出典]」として捉え直す人[誰?]が現れた。 「人類史上類例の無い狩猟採集民」としての縄文人 1970年代以降には更に研究が進展し、それまで動物性食料に依存していたと思われていた縄文人が、実際にはクリなどの堅果類や芋などの根茎類を多く食べ ていたことが明らかとなった。また同時期のヨーロッパの新石器時代人が農耕や牧畜を行っていたとされた[注 1] のに対し、1970年代には縄文人によるヒエ栽培や、縄文後期の水稲を含む稲作の存在が研究者の間でも周知とはなっていなかった。 この結果、縄文人は当時の日本列島の生態系に適合した食料獲得システムを構築し、1万年間の長きに渡って豊かな狩猟採集食文化を維持した、人類史上にも他 に類例の無い人々であったとの見方が登場した。 「海洋民族」としての縄文人 伊豆諸島産の黒曜石が縄文時代やそれ以前に[注 2] 東日本各地で使用されていたことに注目した小田静夫や橋口尚武らの研究により、関東地方の縄文人が縄文早期中葉には内海での漁労に加えて伊豆諸島など外海 へも進出していたことが明らかとなった[50]。 また「縄文土器がバヌアツで表層採取された」というニュース(これについては事実の解釈を巡って議論が続いている[51])にも注目し、縄文人が南太平洋 に進出してポリネシア人の祖先になったという説を唱える者や、エクアドルで縄文土器に似た土器(バルディヴィア土器)が出土したことを理由に、縄文人が南 米大陸に到達していたという説を唱える者さえも出現した[注 3]。 「平和」な縄文人 山口大学と岡山大学の研究グループの人骨調査によると、暴力による死亡率は約1.8%となった。他国や他時代の暴力死亡率と比べると、5分の1以下の水準 である[52]。対照的に弥生時代からは殺害されたとみられる人骨が急増する[53]。 |
・このセクションでは、非常に「自文化中心主義(エスノセントリズム)」的な「縄文人観」が論じられる。1994年以降
の三内丸山遺跡の発掘でブームに火がついた「縄文ブーム」の延長上にある。 ・土偶についてのトンデモ解釈は「『土偶を読む』批判が齎したもの」を参照のこと。 ・岡本太郎は、三内丸山遺跡発見以前の縄文論者であるが、『日 本の伝統』(光文社, 1956年)のなかでこう述べる:縄文の美には、「たんに日本、そして民族にたいしてだけでなく、もっと根源的な、人間にたいする感動と信頼感」がある と。 ・2009年大英博物館で、日本から出展された"The Power of Dogu" では、2か月で7万人の動員をしたそうだ(出典:譽田亜紀子「空前の「縄文ブーム」背後にある日 本人の憂鬱」) ・「アイヌ・コロポックル論争」(コロボックルとも表記) |
| エミシ・エビス・エゾ・アイヌと縄文人 前述のように明治から第二次世界大戦が終わる頃までは、縄文人は日本民族によって日本列島から駆逐されていった先住民と見られていた。こうした見方は必然 的に、古代から近世にかけて日本の支配する領域の北隣に居住していた異民族[注 4]、そしてアイヌを縄文人の直接の末裔と見る説を生み出した。このような縄文人、蝦夷、アイヌを等号で結ぶ見方は、その後の研究の発展によってほぼ否定 され、今日の学界では受け入れられていないが[54]、完全な末裔ではないものの、国立科学博物館が行ったゲノム解析解析によりアイヌ人は縄文人の遺伝子 を特に色濃く残していることも判明している[55]。 近年では、12世紀におけるアイヌ文化の成立をアイヌ民族の成立と見る立場を政治的に不当なものとして糾弾し、古代の北東北からアイヌモシリにかけて広 がっていた擦文文化や続縄文文化の 担い手たちをも「アイヌ」と呼ぶべきであると主張する論者も、少数ながら存在する。例えば平山は山田秀三らが東北地方に アイヌ語地名が多数存在していることを明らかにした研究に言及しつつ、古代の蝦夷(エミシ)と近世のアイヌが同系統の言語を母語としていたことは事実であ り、であるならば古代蝦夷と近世アイヌは同じ民族とするべきであると主張している[56]。小野は12世紀にアイヌモシリでアイヌ文化を生み出した集団 は、11世紀以前にアイヌモシリに居住していた擦文文化人やオホーツク文化人(ニヴ フ系)の直接の子孫であるから、これらは同じ民族と見るべきであると主 張している[57]。 ただ、こうした主張に対しては、エスニック・グループを本質主義的に捉えており、それを構成する人々の形質的特徴や社会的・文化的特徴が長期に渡って不変 であるとの前提に立っていて、現在の人類学・考古学・歴史学・社会学の研究レベルでは通用し難いとの批判がある[58]。 2004年の頭蓋特性の再評価は、アイヌ人がいくつかの特徴において縄文人よりもオホーツク人に似ていて、中間的な性質を持つことを示唆している [59]。また、アイヌの起源が、縄文人に加えてオホーツク人の遺伝的・文化的貢献によるという説もある[60]。 |
・続
縄文文化 ・擦文文化 ・オホーツク文化 ・ニヴフ |
| 縄文人と海 縄文人は基本的には狩猟採集民であったが、その中には海に深く関わっていた人々も存在したことが知られている。 勾玉の分布 遅くともBC5,000年頃(縄文時代中期)には勾玉が作られていたことが判明しており、特に新潟県糸魚川の長者ヶ原遺跡からはヒスイ製勾玉とともにヒス イの工房が発見されており、蛍光X線分析によると青森県の三内丸山遺跡や北海道南部で出土されるヒスイは糸魚川産であることが判明しており、縄文人が海を 渡って広い範囲でお互いに交易をしていたことが考えられている。後年には日本製勾玉は朝鮮半島へも伝播している[61]。 貝類の採集 縄文人が貝類を食糧資源・装飾品の原料として採取するようになったのは縄文早期前半で、代表的な遺跡として横須賀市の夏島貝塚が挙げられる[62]。縄文 早期の半ばには瀬戸内海沿岸や東北地方でも貝塚が形成されるようになる。採取対象は当初は河口等の汽水域に生息するヤマトシジミであったが、やがて内湾干 潟の牡蠣礁で得られるカキや、やはり内湾の軟泥干潟から容易に得られるハイガイなどにその中心は移る[63]。 また、腕輪やペンダントの原材料として採取された貝類もある。特に目立つのが大型の定住性カサガイの一種で岩礁潮間帯低部から採取されるオオツタノハガイ の利用である。オオツタノハガイは主に屋久島やトカラ列島に生息するが、縄文期には、特に縄文後期・晩期を中心に、関東全域から北は北海道の有珠10遺跡 でも出土している。これについて、原材料となったオオツタノハガイは南九州から運ばれたという説と、三宅島以南の伊豆諸島にも生息域があったのではないか との説が対立している[64]。 オオツタノハガイの他には暖流域の浅海から得られるタカラガイの一種ハチジョウタカラガイも広く利用された。 伊豆諸島への進出 前述のように、先史時代の日本列島住民が今日の伊豆諸島に進出したのは旧石器時代である。しかし、縄文期の遺跡に限ると最も早いものでも縄文早期の半ばの ものとなる。この時期の遺跡としては伊豆大島下高洞遺跡、神津島せんき遺跡、三宅島の釜ノ尻遺跡などがある[65]。 縄文前期の末には黒潮の本流を越えた[注 5]。縄文人が八丈島に進出し[注 6]、倉輪遺跡からは関東、南東北、中部、関西地域の土器が発見されている。 九州島と南島・朝鮮半島間の交流 縄文前期には九州島[注 7] を中心として轟式土器と呼ばれる土器が広く使用されるようになった。轟式土器は九州島周辺の他、種子島や屋久島、朝鮮半島南部にも分布しており、これらの 島々・半島間を航行した縄文人集団が存在したことをうかがわせる。日本列島周辺や南西諸島周辺、朝鮮半島周辺の島々は国ができる以前からこれらの海域を行 き来する海洋民族によって既知だったと推測される。 また轟式に続いて登場した曽畑式土器も、奄美大島の高又遺跡、沖縄島の読谷村渡具知東原遺跡、朝鮮半島の慶尚南道にある釜山市の東三洞貝塚などから発見さ れている[66]。縄文人が黒潮本流を越えた例としては、この曽畑式土器を持った集団による縄文前期の九州島・奄美大島間の航海が最も古く、関東における 三宅島・八丈島間の航海よりおよそ800年早いものであるとされている[67]。東三洞貝塚では大量の縄文土器と北松浦半島産黒曜石が出土しており、縄文 人がやってきた確かな証拠といえる。なお、朝鮮半島では銛や鏃に最適な黒曜石が産出されない[68]。 長浜浩明は、大阪市立東洋陶磁美術館名誉館長の伊藤郁太郎の見解として、「1969年から1971年にかけて東三洞貝塚の下層から尖底・円底無文土器が発 見された。これらの中には北松浦半島の泉福寺洞穴や福井洞穴などから発見された隆起線文土器と類似する土器、同じく北松浦半島の黒曜石と大形石斧(佐世保 市相浦の門前遣跡[69] 製)も含まれていた。その他、慶尚南道真岩里や咸鏡北道西浦項貝塚などからも発見されている」と紹介し、縄文人は7000年前から無人の朝鮮半島へ渡り、 半島北部まで進出していた、と述べている[70]。また、朝鮮半島南部の煙台島貝塚から発見された古人骨(紀元前4000年)は縄文人の特徴と多くの点で 一致しており[71]、現代韓国人とは似ても似つかぬ形態であり、最初に半島に住み始めた人々は日本からやって来た縄文人だったという考古学からの推論 が、形態人類学によって裏付けられたとしている[72]。また長浜は、朝鮮半島の櫛文土器文化の土器は九州の曽畑式土器(縄文前期)そのものであると述べ ている[73][74]。 縄文人の用いた舟艇 これまでに出土した事例に見る限り、縄文人が航海に用いたのは一本の丸太を刳り抜いた丸木舟であったと考えられている[注 8]。帆柱の跡やオール受けの跡は検出されていないため、(カイトセイリングのように帆柱を用いない形式での帆走を行った可能性は否定出来ないまでも)基 本的にはパドリングによる推進であった可能性が高い。 船体の断面は関東地方出土の丸木舟を見る限りでは半月型[注 9] あるいは三日月型であり、弥生時代以降の凹型断面の丸木舟とは異なる特徴を示している。船体長は最大で残存長7メートルから8メートルのものまであるが (例えば千葉県香取郡多古町島(七升)出土の縄文前期のものは残存長7.45メートル、残存幅0.7メートル)、小さいものでは4メートル以下のものも多 数出土している。 材はアカマツやクロマツ、カラマツ、カヤ、ケヤキ、ムクノキ、クスノキなどの例がある[75]。 なお、1982年には松江市内の小中学校の教師の有志5名により、「からむしII世」と名付けられた丸木舟による黒曜石の運搬実験が行われ、隠岐の宮尾遺 跡から本州の松江市美保関町の七類港まで15キロの黒曜石を1日で運搬することに成功している[76]。 |
|
| 言語 縄文人の言語については明らかでない。日本語のほか、人類学的類似性が高いとされるアイヌ(アイヌ語)との関連を指摘する見方[77] もあるが、推測の域を出ない。過去の言語は文字がなければ検証不可能であり、「縄文語」の解明は、その試み[78] は存在するものの、極めて難しいと言わざるを得ない。日本列島における縄文時代の言語は、共時的にも通時的にも複数存在したと考えるのが自然である。 |
|
| 日本人 弥生人 古墳人 倭人 先住民族 ハプログループD (Y染色体) ハプログループD-M64.1 (Y染色体) ハプログループC1a1 (Y染色体) ハプログループM7a (mtDNA) ハプログループF (mtDNA) ハプログループB (mtDNA) 縄文時代 縄文時代の遺跡一覧 後期旧石器時代 勾玉 徐福 山崎三四造 - 縄文人の生活を20年間にわたり再現・実践した。 古代北部東アジア人 古代北ユーラシア人 |
|
| 日本語ウィキペディア「縄文人」https://x.gd/3W2p6 |
リ ンク
文 献
そ の他の情報
Copyleft, CC, Mitzub'ixi Quq Chi'j, 1996-2099
![]()
☆
 ☆
☆
{kind=link}