「日本人起源論」の系譜
So
called, the Origin of Japanese..
| このページの目的は、いわゆる「日本人起源論」のなかに、
(1)歴史的
時間概念の「脱臼」あるいは土着神話学、(2)人類学理論における「伝播論」
の影
響、(3)ナショナリズムにもとづく「ナショナル・アイデンティティの探求」、(4)自己表象と他者表象の表現のなか
にみられる「レイシズム(人種主義)」、(5)日
本人という「ナショナリティへの固執」がみられることを検討することです。 ★「そもそも人間をグループ分けする必要があるところに問題があるのだ。たとえ分割することが、異なるかもしれないごく小さなゲノムの断片を拡大しなけれ ばればならないことを意味し、それですら平均値の話だとしても、人間を分割することが問題なのだ。分割し、人々を異なったものとして扱うこの必要性こそ、 人種を生みだした過程なのだ」(サイニー2020:185) |
★【再掲】このページの目的は、いわゆる「日本人起源論」のなかに、(1)歴史的時間概念の 「脱臼」 あるいは土着神話学、(2)人類学理論における「伝播論」の影響、(3)ナショナリズムにもとづく「アイデンティティの探求」、(4)自己表象と他者表象 の表現のなかにみられる「レイシズム(人種主義)」、(5)日本人という「ナショナリティへの固執」がみられることを検討することです。
★片山一道は言う、701年大宝律令後の律令国家が8世紀以降まで「日本」なるもの観念は形 成 されなかったので、それ以前の「日本人」とは、日本列島に居住する人を、便宜的にそう呼ぶしかない(片山 2015:16-17)。
★「第二次世界大戦後、長谷部言人と鈴木尚は、日本人の起源は弥生時代の渡来人ではなく、縄 文 時代の人々であると主張した。しかし、埴原和郎は1984年に新 しい人種混血説を、1991年には「二重構造モデル」を発表した。埴 原氏によれば、現代日本人の血統は、旧石器時代に日本列島に移住してきた縄文人に始ま り、弥生時代(紀元前300年)には東アジアから日本への第二の移住の波が押し寄せた。新石器時代に人口が拡大した後、弥生時代のある時期にこれらの新人 が日本列島にたどり着いた。その結果、九州、四国、本州南部の島嶼部では狩猟採集民の交替が一般的であったが、沖縄や北海道の離島では優勢ではなく、琉球 人とアイヌ人は混在した特徴を示している。マーク・J・ハドソンは、日本人の主要な民族像は紀元前400年から紀元後1,200年にかけて生物学的、言語 学的に形成されたと主張している。現在最も有力な説は、現在の日本人は弥生時代の稲作農耕民と縄文時代の諸民族の両方から形成されたというものである。し かし、最近の研究では、縄文人には当初考えられていたよりも多くの民族的多様性があったという説や、日本人は2つの古代集団だけでなく、3つの古代集団か らの重要な遺伝的特徴を持っているという説もある。」英語ウィキペディアJapanese peopleより).[→「日本人起源論のな かにみられる人種主義」]
★コロニアリズムと日本人起源論の歴史的関係(→今後は「日本人起源論のなかにみられる人種主義」で改善し ます)
1)1940年代以前:日本人の渡来起源説が定番 だったが、第二次大戦の激化とともに皇民化政策がすすみ、天皇を頂点とする「純粋な人種」としての日本人のイデオロギー的要請が増えてきた。しかし、それ を自然人類学的に「論証」した研究者はいなかった。
2)1940年代後半、長谷部言人などは、混血がなかったことをこの時期に主張するようになる。長谷部言人「『日 本民族の成立』(昭和24年)では、前期洪積世以降の日本列島住民の転変を、身体と文化の両面から考察し、縄文人と古墳時代人との体質的差異は、狩猟採集 経済を基盤とする石器時代の生活から、水田農耕に依存する金属器時代の生活への転換が、租借筋、下肢筋の弱体化を招く結果を生じたと解釈し、弥生式時代お よびその後においても、日本人の体質を一変させるほどの混血はおこらず、日本人は石器時代から現代にいたるまで遺伝的に連続した集団である」と主張。
3)1950年代から60年代にかけて鈴木尚は、 「長谷部言人らの流れをくむと共に自らの長年の研究、なかんずく縄文時代から弥生時代へと移行する時期の人骨の詳細な調査検討に基づき、縄文時代人が弥生 文化の流入に伴う生活環境の変化のため、いわゆる小進化によって弥生時代人に変わったという「変形説」を主張した」。この流れは、1970年代まで(日本 人起源論の)支配的パラダイムとなる。
4)他方、1950 年代から 1970 年代には、台北とソウルの日帝の旧植民地の自然人類学者は、金関丈夫の渡来説を展開し混血渡来説を多く支持するようになる。彼らの見解によると、日本人は 異種混交の「民族」(=意識的に「純粋な単一の人種」を否定する)であり、金関がリー ダーシップをとった、山口県土井ヶ浜遺跡(1950〜)の発掘 し、渡来説を主張するようになる。
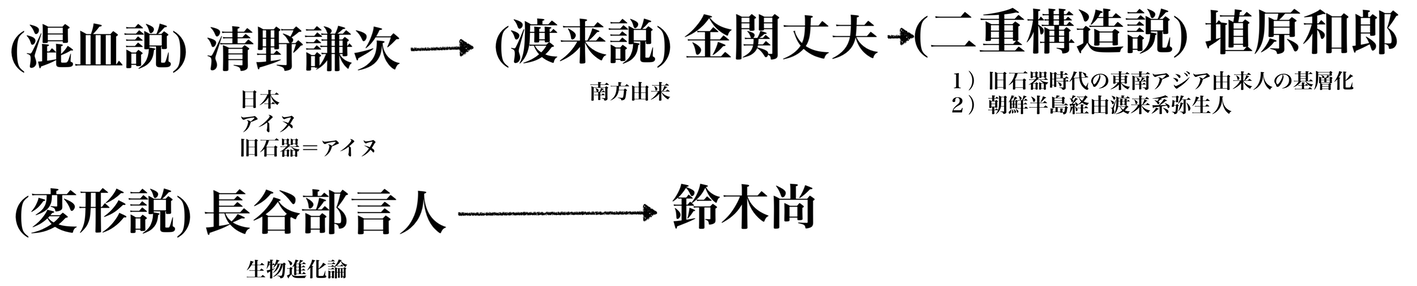
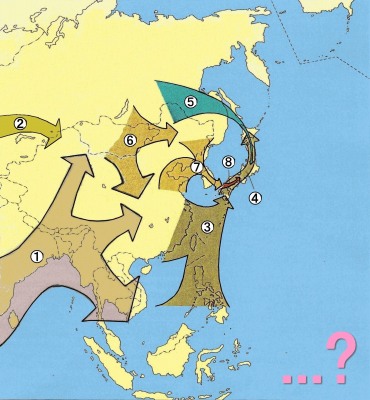
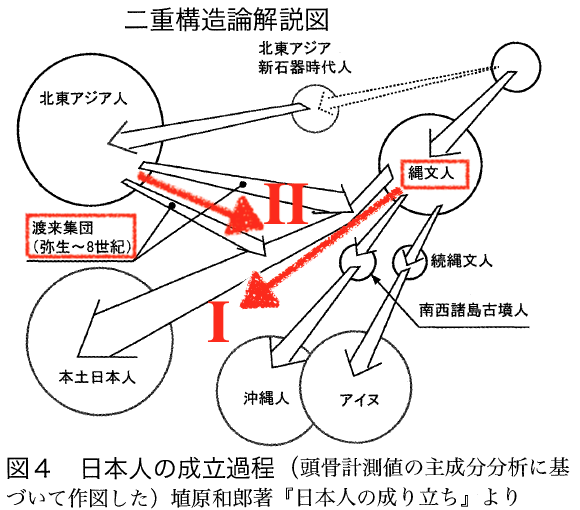
5)1980年代から90年代にかけて、それまでの 計測研究を中心とする自然人類学研究に、遺伝学的研究が加わり、遺伝的形質の類似性と多様性があることがわかり、それが埴原和郎の1991年の「二重構造モデル」につながる。これは、東南アジア起源の 縄文人と いう基層集団の上に、弥生時代以降、北東アジア起源の渡来系集団が覆いかぶさるように分布して混血することにより現代日本人が形成された。渡来系集団は、 北部九州及び山口県地方を中心として日本列島に拡散したので、混血の程度によって、アイヌ、本土人、琉球人の3集団の違いが生じた、と説明するものである (→「二重構造モデル」)。
6)二重構造モデルが、本土日本人の混血を主張するために、単一人種=民族と
しての日本人説が払拭されたわけではない。埴原のこのモデルでは、弥生から8世紀の後には、混血がおわり、「日本人種」は一千年以上安定した集団になるわ
けであるし、また「日本人」としての、沖縄人やアイヌは、縄文人として「古代的」な特徴を保持し、それ自体は、近代化以降における、アイヌ人や沖縄人への
(形質的差異にもとづく)「人種的偏見」の原因になってきたことは、歴史社会学的によく知られる事実である。
★土肥直美(2011:36)によると、起源論は3つに分けられる;1)置換説(交代説)、
2)混血説(渡来説、渡来混血説)、3)変形説(小進化説、連続説、移行説)
★池田次郎(1998)による年代区分である(→「起 源論各説」より)。
★表象のなかのレイシズム(→「日本人の起源における二重構造モデルの攻防」を参照)
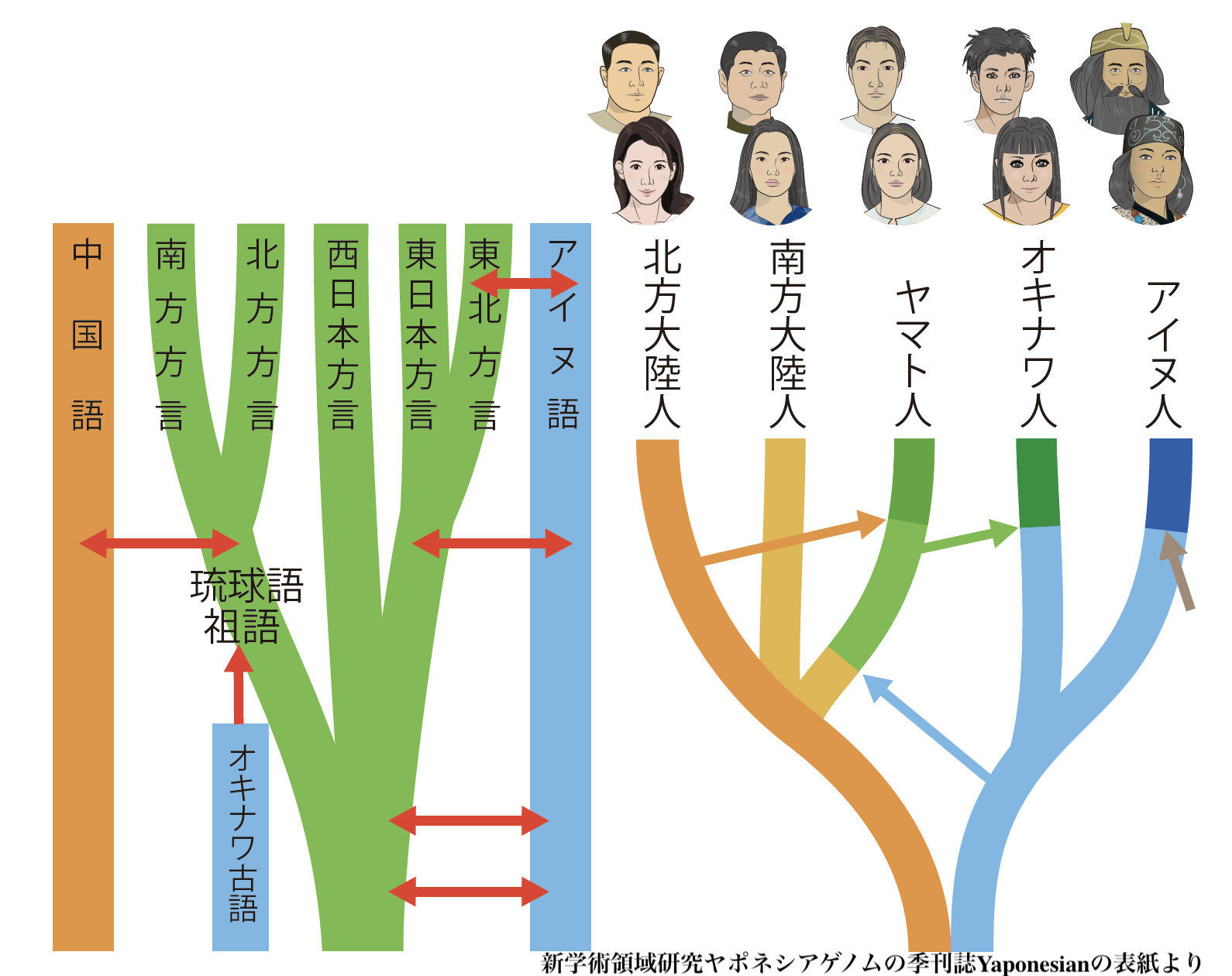
「新学術領域研究ヤ ポネシアゲノム」研究は、はたして、科学人種主義の特徴を持つのか?(→「日本人起源論のなかにみられる人種主義」)
| 第1期(江戸時代末~大正時代末) ・人種交替説:先住民(石器時代人)と渡来人(金属 器時代人)の交替 ・プレ・アイヌ説(モース)、コロボックル説(坪井 正五郎) ・アイヌ説(シーボルト、小金井良精)、固有日本人 説(鳥居龍蔵) |
|
| 第2期(昭和初年~昭和30年ごろ) ・原日本人説:縄文人は原日本人あるいは先史日本人 ・混血説(清野謙次)―渡来人の影響を想定;津雲貝塚など千体の縄文人骨の統計的研究をもとに主張。縄文人は現日本人、混血を経て日本人に、アイヌは縄文 の形質を残す。 ・変形説(長谷部言人)―文化的影響で縄文人から弥 生人へと変化し、現代日本人に変化していく。 |
・須田昭義(1950)「人
類学からみた琉球人」『民族学研究』15(2):109-116.→琉球人は日本人の一地方型にすぎないと報告(1950:113-114) ・許鴻梁(木偏に梁)『琉球人頭骨の人類学的研究』(1950)「琉球人頭骨の人類学的研究」『国立台湾大学解剖学研究室論文集』2(1948)——琉球 は南方集団と比較的近いがアイヌとは遠い関係にある。 |
| 第3期(昭和30年代~昭和50年代) ・渡来説(金関丈夫)―地域=九州を限定して渡来を認める。縄文との混血が進む。 ・小進化説(鈴木尚)―変形説を強化。生活によって縄文から弥生的な変化は可能と考える。 |
|
| 第4期(昭和50年代~) ・渡来説が復活し、定着 ・縄文人と渡来系弥生人の原卿が議論の焦点となる ・埴原和郎「二重構造」説は、基本的に「混血説」 |
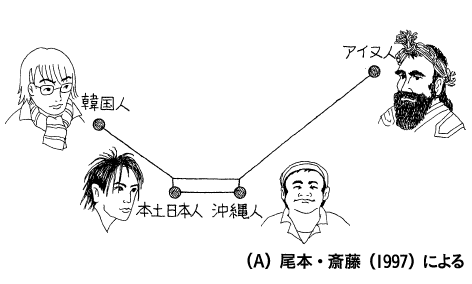
・池田次郎と多賀谷昭(1976)は、アイヌと琉球に共通の特徴は認め
られるが、同時に日本人集団とも類似すると結論 ・埴原和郎「二重構造」説は、基本的に「混血説」であるが、日本本土中心的にみるのではなく、日本本土は渡来人からの混血が強く(混血の機会が二度あった ので「二重構造」という)、結果的に、アイヌと琉球に古い特徴が残るという、ベルツの同祖論の再演と理解することも可能である。 ・ただし土肥によると、アイヌ・琉球同系論 についても、最近、疑問が投げかけられている(→「日本人の起源における二重構造モデルの挫折」) |
【再掲】
第1期(江戸時代末~大正時代末)
・人種交替説:先住民(石器時代人)と渡来人(金属 器時代人)の交替
・プレ・アイヌ説(モース)、コロボックル説(坪井 正五郎)
・アイヌ説(シーボルト、小金井良精)、固有日本人
説(鳥居龍蔵)
第2期(昭和初年~昭和30年ごろ)
・原日本人説:縄文人は原日本人あるいは先史日本人
・混血説(清野謙次)―渡来人の影響を想定;津雲貝 塚など千体の縄文人骨の統計的研究をもとに主張。縄文人は現日本人、混血を経て日本人に、アイヌは縄文の形質を残す。
・変形説(長谷部言人)―文化的影響で縄文人から弥
生人へと変化し、現代日本人に変化していく。
第3期(昭和30年代~昭和50年代)
・渡来説(金関丈夫)―地域=九州を限定して渡来を 認める。縄文との混血が進む。
・小進化説(鈴木尚)―変形説を強化。生活によって
縄文から弥生的な変化は可能と考える。
第4期(昭和50年代~)
・渡来説が復活し、定着
・縄文人と渡来系弥生人の原卿が議論の焦点となる
上掲「起 源論各説」のHP解説より:「日本人の起源をめぐる論争の歴史は古く、すでに江戸時代から、新井白石や木内石亭らによる著述が世に問われていた。 本格的な研究は、幕末から明治始めにかけて来日した外国人研究者によって開始され、以後「日本人の起源」は、日本人類学会の最大の研究テーマとして、現在 に至るまで熱い論争が続けられている。とくに昭和30年代以降の研究進展には、金関丈夫、永井昌文らによって収集、保管されてきた、弥生人骨を柱とする九 州大学の人骨資料が決定的な役割を果たした。現在もなお、国内はもとより海外からも多数の研究者が訪れ、多くの研究成果が発表され続けている。」
●二重構造モデル(詳しくは「いわゆる「二重構造モデル」:日本人起源論の系譜仮説」)
日本人の起源における 「二重構造モデル」とはざっと次のようなものである;「現代日本人の形成に関する埴原和郎・東京大学名誉教授の仮説。すなわち、東南アジア起源の縄文人と いう基層集団の上に、弥生時代以降、北東アジア起源の渡来系集団が覆いかぶさるように分布して混血することにより現代日本人が形成された。渡来系集団は、 北部九州及び山口県地方を中心として日本列島に拡散したので、混血の程度によって、アイヌ、本土人、琉球人の3集団の違いが生じた。この仮説は大筋では受 け入れられているが、基層集団の起源が北東アジアではないかとの意見も強い」馬場悠 男, 2007)。
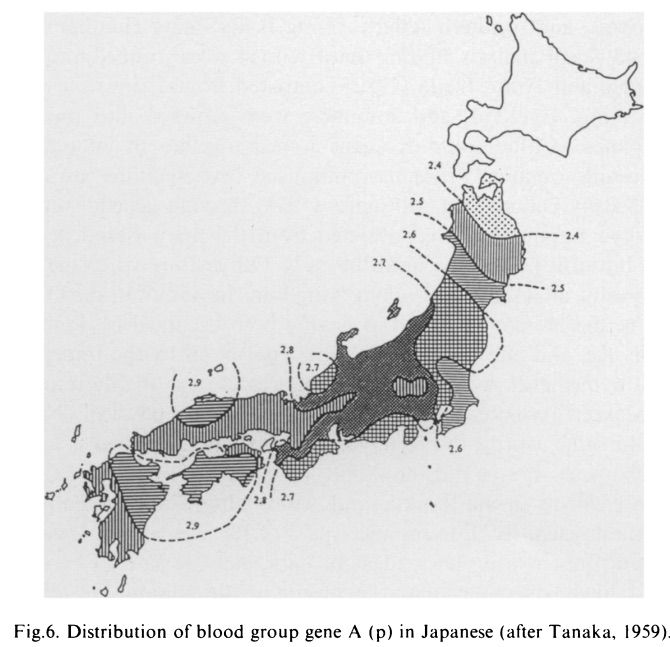
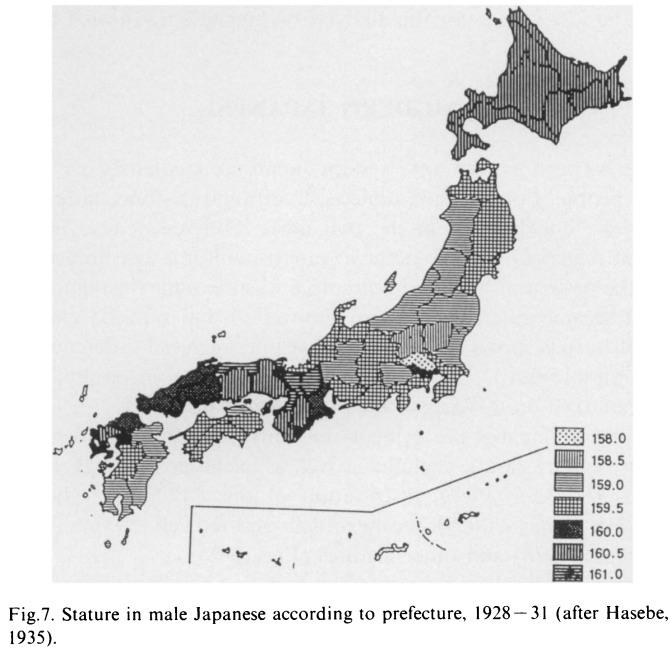
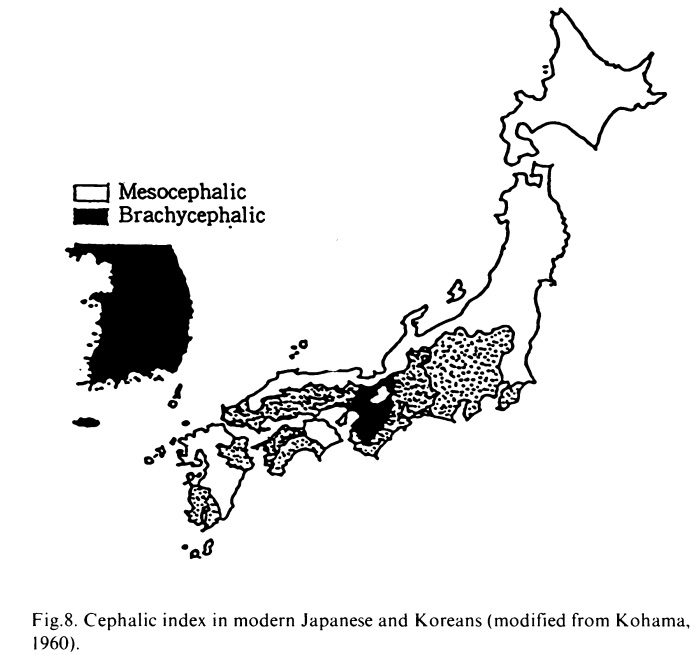
●沖縄からみた「二重構造モデル」批判→「日本人の起源における二重構造モデルの挫折」を参照。
「沖縄に関しては、この他に、かなり一般的になって いるアイヌ・琉球同系論 についても、最近、疑問が投げかけられています。東北大学の解剖の教授をしている百々 幸雄さんは、私たちと同じ骨を研究対象にしていますが、私たちが計測値を比較するとい う手法を用いるのに対して、百々さんは計測できない特徴を研究対象にしています。それ は人の個性みたいなもので、神経や血管の通る穴が―つの人もいれば二つの人もいる。頭 の骨にはそういう非計測的形質といわれる特徴がたくさんあるんです。いろいろな特徴の 出現頻度をアイヌと沖縄と本土の日本人で比較すると、これまで言われているように、琉 球とアイヌが近縁という結果にはならなかったんです。それで琉球=沖縄人とアイヌが 同 系であると言えるかどうかの見直しをしなければならなくなりました。/ 私もその共同研究に参加しています。この共同研究では計測値についても顔面平坦度と いう新しい計測項目を取り人れて、詳しい分析を行いましたが、その結果はアイヌと琉球 =沖縄人はかなり違うというものでした。一般的に琉球=沖縄の人はほりの深い顔立ちを していると言われるんですが、骨で見る限りでは非常に平坦な顔立ちという結果になり、 私たち自身が大変驚いてしまいました。琉球=沖縄、アイヌ、本土日本というのがそれ ぞ れに近い関係ではあるけれども、独自の特徴を持っているという結果です。骨はそれぞれ の地域の歴史をかなり正直に反映しているのではないでしょうか。人の 動きはもっと複雑 で、もっとダイナミックなものだったと思います」(安里・土肥 2011:58-59)。
……
……
★篠田謙一「第6章日本列島集団の起源:本土・琉球列島・北海道」『人類の起源』中公新書、 中央公論新社、pp.195-231、2022年
|
1. 日本人のルーツ
2. 琉球列島集団
3. 北海道集団
|
1. 日本人のルーツ
■二重構造モデル
1)縄文人と弥生人の形態の違い;2)北海道アイヌ集団、琉球集団、本土日本人の、「異なる 形態」の集団が存在する
二重構造:1)旧石器時代に東南アジアから北上して、縄文人になる。2)大陸を北上した集団 は寒冷地で適応して、東北アジアの新石器人となる。3)弥生時代になると朝鮮半島経由で、東北アジア人が九州に稲作をもたらす。後者が「渡来系弥生人」で ある。縄文と弥生の形態の違いは、集団の違いによる(196)
・古代ゲノム解析の結果、大陸沿岸部と内陸部を北上した2系統のアジア人がいる。大陸北部で
の寒冷地適応はあった可能性。最終氷期最寒期(1,9000年前)に田園洞遺跡の人骨に近い集団に、入れ替わる。EDARの特異系頻度が増える。370A
は寒冷地適応の形質に関連する。
■二重構造モデルの視点
・渡来人と縄文人の混血はすすむが、北海道と琉球は、稲作ができない(北海道)と2000年 遅れた琉球では、縄文的形態が残った。
・二重構造モデルは、「遅れた」北海道と琉球という時間的モデルを、空間的な分布に関連づけ ている点で留意が必要だ。ひとつの視点で(=日本という視点)で南北三千キロの日本列島の人間の成り立ちを説明できない?可能性。
■ゲノム解析による地域別比較
・本土のゲノムはよく似ているが、近畿・四国と、九州、東北のあいだの3つの集団の違いが SNP解析で見えてくる(ただし198ページの図はグラデーションマップの形をとる)。畿内では渡来系の影響がつよく、周辺では縄文系が残る。二重構造 は、この類推で、北海道と琉球は縄文と推測する(199)
・集団の形成は、地域的であるのに、二重構造は日本全体を一つのモデルで説明しようとするの で、無理がある。「地域の歴史の総和が列島集団であるという発想」が ない(200)。北海道(アイヌ)と本土と琉球は、それぞれ、個別の形成をしている(つまり、二重構造論は「プロクルステスの寝台」だというのだ)。
・その理由を篠田は2つ指摘する;1)人骨形態の研究の限界、2)日本人や縄文人という発想 が「地域の歴史の総和が列島集団であるという発想」に結びつかない (200)。篠田の結論は、地域別に集団のなり方を考えるべきだ、ということになる。
■旧石器時代
・6万年前。遺跡はあるが人骨はみつからない。唯一沖縄のみ。
・ハプロタイプとは、ある程度遡ると祖先が同じになるグループである。ハプロタイプとは "haploid genotype"(半数体の遺伝子型)の略語。大まかにL=アフリカ集団、NとR=ヨーロッパ集団、M=アジア集団(76)
・唯一の旧石器人である港川人は、Mに属するが6万年に発生して、港川人の2万年のあいだに 変異がもっとあってもいいのだが、それが見つからないために、港川人は現在の「日本人」には繋がらず、消滅した(絶滅?)可能性がある(201)
・現代の沖縄のゲノム解析よって、その祖先は1万5千年よりも遡らないので、2万年前の港川
人とのつながりはない可能性が高い。ただし、ミトコンドリアゲノムは母系の祖先だけを調べることができる(201)。
■縄文人のミトコンド リア系統
・これまでに100体のミトコンドリアDNA解析がおこなわれた。
・縄文人のハプログループはM7aとN9bで、日本国内のみ。双方とも、成立は3万〜2万年 前なので、縄文よりも前に、日本列島に入ってきて、大陸の先祖は消滅してしまった(=篠田は絶滅ということばは使わない)(203)
・N9bは、東日本から北海道、M7aは、西日本から琉球列島で多数派をしめる。M7aは大 陸からやってきて、東のほうにすすんでいった。琉球列島と九州との交流1万年前に分岐し、それ以降はまじることがなかった(203-204)
・N9bは、複雑で、移動のプロセスを説明できていない(204)。九州縄文人にN9がある ので、N9は、朝鮮半島と、沿海州の北回りで、日本にやってきた可能性があり、M7aのような単純な説明ができない(204)
■縄文人の地域
・縄文人は旧石器時代にさまざまな地域からはいってきた集団である。(篠田は記載していない が、人口はそう多くなかった)。また、縄文人というエスニックアイデンティティもなかったろう。縄文人をひとつの集団として理解することの限界 (205)。人口分布は、東高西低。現代日本人は、M7aが7.5%、N9bは2.1%(205)
■縄文人の核ゲノム解析
・2016年三貫地遺跡の縄文人の核ゲノム分析がおこなわれる(206):女性北方系、現代
のアイヌや沖縄と形態的に似ている遺伝子。婚姻のネットワークは小規模で、だが両親のあいだには三親等の血縁関係はなし(207)
■縄文人と東アジア現代人との関係
・東アジアの集団との比較で考える(208)。縄文人は、大陸集団より早く分岐して、長期間 独立して独自の遺伝的特徴を獲得していった(208)。現代日本人との比較では、北東アジアの集団が日本列島に入って、縄文系と混合したことがわかる (208)。朝鮮半島の集団にも、その基層にある縄文につながる遺伝子がある(→縄文の祖先の大陸起源の根拠)。
・縄文の遺伝的特異性(209):縄文の近縁性探していくと、アイヌ、琉球、本土のほかに、 台湾、韓国、沿海州の先住民とも類縁関係を確認できる(209-210)→東南アジアから北上してきた集団のなかで、沿岸に住んでいた人たちが縄文の母体になった可能性 (210)
・ハプログループの解析により、縄文人は従来考えれるよりも、多様性をもった集団である
(210)。
■弥生時代
・弥生時代定義は、稲作をはじめたことによる定義(210)——北部九州で3000年前。弥 生時代は、弥生式土器、水田稲作、金属器の使用(211)。農耕と金属器は外来から。
・縄文のハプログループは単純だが、現代に日本人のそれは多様であり、それは渡来した人々の 影響である(212)
・日本本土人は、渡来した人たちによって形成されている(212):弥生時代は、渡来人、縄
文人、そしてその両方の「混血」からなる。九州でもその3つの集団を想定できる→図6-5(p.213)
■弥生人のゲノム
・朝鮮半島でも沿岸部では(=現代日本人と共通の)「縄文的な遺伝要素」をもった人が出土す る。この縄文的とは、東アジアの沿岸部の旧石器時代集団の遺伝的要素のことである。
・現代韓国人が、中国東北部の集団と遺伝的に「区別」できるのも、この縄文的な要素によるも
のか?(215-216)
■列島集団と大陸系集団
・弥生時代の朝鮮半島南部の人々が、現代日本人とおなじような遺伝子組成をもつことは、縄文 人と弥生人がまったく別の遺伝子をもつという考え方に再考をせまる(216)
・朝鮮半島古代集団、渡来集団、西遼河の新石器雑穀農耕民の3者の遺伝的連続性がある。あわ
せて山東半島の農耕民が、朝鮮半島ならびに日本に稲作農耕を伝える。(図6-6)
■弥生人の核ゲノム解析
・西北九州の弥生人も、(形態的特徴から)縄文人の子孫と考えるよりも、渡来人との混血と考 えるほうが適切——すくなくともゲノムではそうなった(218)
・「ゲノムで見ていくと、弥生人には集団としての実態があるわけではない」(219)→「集 団としての実態」は確固とした遺伝子集団としての「レイス」を想定されるが、縄文系もふくめて、多様な遺伝子の存在があり、むしろ、時代をへるうちに、弥 生人としてのゲノムの均一化(→言い方をかえると「レイスとしての日本人」の形成)があったと指摘(219)——篠田は「レイス」ないしはそれに相当する 「人種」の用語は一切つかわない。だが「均一化」として遺伝的多様性が少なくなることを、一定の集団的特質性としてみていることは確かである。
・渡来系集団は、混血を通して「縄文人の遺伝子を次々に取り込んでいった」(219)
・弥生中期にも、大陸からの渡来を想定しないと、現代日本人の遺伝的特徴を説明できない (220)
・「現代日本人の形成
のシナリオは弥生から古墳時代における大陸からの集団をいっそう考慮したものになるでしょう」(220)
2. 琉球列島集団
■琉球列島集団の成立
・旧石器時代のゲノム研究は港川人のミトコンドリアDNAのみ(221)
・縄文時代(編年は琉球では異なる)の人骨形態は、本土の縄文人とは異なるものの、ミトコン ドリアDNAでは九州の縄文人と共通(222)
・12世紀ごろからはじまるグスク時代に人口急増(222)
■琉球列島集団のミトコンドリアDNA
・縄文時代は、本土の縄文人のハプログループのM7aとN9bしか出現しないが、弥生平安時 代(貝塚後期)になると、少数ながら本土由来のハプログループがみつかる(222)
・グスク時代初期に奄美で南九州からの農耕民の移住の考古学的証拠がみつかる(223)
・現代の琉球列島集団に縄文人由来のゲノムが30%残っているので、島外からの影響は本土に くらべると少ない(223-224)
・琉球大学が大規模なゲノム研究をおこなっていて、今後の研究が期待される(224)
3. 北海道集団
■北海道集団の成立
・オホーツク文化(→オ ホーツク文化とそれを担った人びと)※オホーツク文化は、二重構造モデルでは、アイヌ文化に影響を与えず姿を滅ぼし、アイヌは縄文直系の子孫とい われてきた(225)
・しかし、ミトコンドリアDNAの解説からオホーツク文化人の影響をうけているとわかってきた(226)
・平取のデータでは、アイヌは本土日本人とのあいだで「混血」がすすんでいることが明らかに なった(227)
・礼文島での研究では、カムチャッカ半島集団から2,000年前に、沿海州から1,500年 前に遺伝子の流入。これがオホーツク文化人のアイヌへの影響と考えられる(227)
・ただし、続縄文か ら擦文文化にかけての時代(続縄文→オホーツク→擦文)の骨がみつかっていな い(227)
・本土日本人は渡来人の末裔、アイヌ人は北海道の縄文の末裔(228)——ただし、アイヌは 北方からの遺伝子も受け取る。
・現代沖縄人に、台湾より南方から遺伝的影響はみられない。
・二重構造モデルは日本を中心にして考えて、周辺(この場合は、北海道アイヌと琉球人)の遺 伝的組成を考えないので、周辺の民族の遺伝的組成を説明することができない(228)
・だから北海道を中心にする「集団の変遷のシナリオをつくる必要性がある」と結論(228) →《そのためのアイヌ遺骨のゲノム解析は欠かせない》という論法。
※コラム4:倭国大乱を示す人骨の証拠
★国立科学博物館・溝口優司による「更 新世から縄文・弥生期にかけての日本人の変遷に関する総合的研究」の説明(カラー図)
「ア フリカで現代人(ホモ・サピエンス)にまで進化した集団の一部が、約6〜3万年前、東南アジアや東アジアへやって来て、その地の後期更新世人類となった (1,2)。次いで、更新世の終わり頃、約1万年前までに、東アジアや東南アジアの後期更新世人類が日本列島に到達し、その子孫が日本列島全体に広がって 縄文時代人となった(3,4)。同じく更新世の終わり頃、北方からも日本列島へ移住があり、それが縄文時代人の形質に地理的勾配を生じさせたかもしれない (5)。他方、後期更新世のいつの頃か、もとはと言えば縄文時代人などとルーツは同じだがシベリアや北アジアで寒冷地適応した集団が東進南下し、少なくと も6000年前までには中国北東部、朝鮮、中国の黄河流域・江南地域などに分布して、その地域の新石器時代人となった(6)。そして、 縄文時代の終わり頃、中国北東部から江南地域にかけて住んでいた人びとの一部が朝鮮半島経由で西日本に渡来し、先住の縄文時代人と一部混血しながら、広く 日本列島に拡散した(7,8)。これが弥生時代以降の本土日本人の祖先である。この渡来民は沖縄の人びとにも遺伝的影響を与えたが、アイヌの人びとにはあ まり影響しなかった。アイヌは縄文時代人が少しずつ変化して生じた人びとである」
出典:https://www.kahaku.go.jp/research/department/anthropology/report02/s14.html
リンク
リンク
文献
その他の情報


++
Copyleft, CC, Mitzub'ixi Quq Chi'j, 1996-2099
![]()
☆
 ☆
☆